Fenofibrate, a peroxisome proliferator-activated receptor alpha-agonist, blocks lipopolysaccharide-induced inflammatory pathways in mouse liver
- Affiliations
-
- 1Department of Surgery, School of Medicine, Wonkwang University, Iksan, Korea. furufuru79@naver.com
- KMID: 1472524
- DOI: http://doi.org/10.14701/kjhbps.2013.17.3.89
Abstract
- BACKGROUNDS/AIMS
During the acute phase response, cytokines induce marked alterations in lipid metabolism including an increase in serum triglyceride levels and a decrease in hepatic fatty acid oxidation, in bile acid synthesis, and in high-density lipoprotein levels.
METHODS
Peroxisome proliferator-activated receptors (PPARs: PPARalpha, beta/delta, and gamma) regulate fatty acid metabolism, glucose homeostasis, cell proliferation, differentiation and inflammation. Proinflammatory profiles including tumor necrosis factor alpha (TNF-alpha), interleukin-1beta (IL-1beta), and interleukin-6 (IL-6) are the important pathological factors in inflammatory responses during the pathological progression of the acute phase response. Lipopolysaccarides (LPS) induced the expression of TNF-alpha, IL-1beta, and IL-6. LPS-induced inflammation decrease the expression of peroxisome proliferator-activated receptor alpha (PPARalpha), PPARbeta/delta, PPARgamma, and coactivators PPARgamma co-activator 1 alpha (PGC-1alpha), PGC-1beta messenger RNA (mRNA) in the liver of Balb/c mouse. In addition, LPS-induced inflammation diminishes the protein level of PPARalpha, PPARbeta/delta, and PPARgamma. Proinflammatory cytokines including TNFalpha, IL-1beta, and IL-6 are the principal reducer of PPARs. However, the knockout mouse model against TNFalpha and IL-6 does not block decrease of PPARs in serum and liver. The mice were pretreated with fenofibrate at 100 mg/kg for 2 days.
RESULTS
These treatment protocols increased the amount of PPARs mRNA in the liver. Fenofibrate inhibited LPS-induced TNF-alpha, IL-1beta, and IL-6 production in the serum and liver. Similar results were obtained when human hepatoma HepG2 cells exposed to LPS were co-incubated with fenofibrate. LPS-treated HepG2 cells decreased expression of IkappaB. Moreover, activation of PPARs abrogated LPS-induced degradation of IkappaB, thus suppressing LPS-induced NF-kappaB activities.
CONCLUSIONS
Therefore, fenofibrate decreases the expression and secretion of TNF-alpha, IL-1beta, and IL-6 via the NF-kappaB signaling pathway, thus serving as therapeutic targets to attenuate inflammation that is involved in hepatic pathological progression.
MeSH Terms
-
Animals
Bile
Carcinoma, Hepatocellular
Cell Proliferation
Clinical Protocols
Cytokines
Fenofibrate
Glucose
Hep G2 Cells
Homeostasis
Humans
Inflammation
Interleukin-1beta
Interleukin-6
Lipid Metabolism
Lipoproteins
Liver
Mice
Mice, Knockout
NF-kappa B
Peroxisome Proliferator-Activated Receptors
Peroxisomes
PPAR alpha
PPAR-beta
PPAR delta
PPAR gamma
RNA, Messenger
Tumor Necrosis Factor-alpha
Cytokines
Fenofibrate
Glucose
Interleukin-1beta
Interleukin-6
Lipoproteins
NF-kappa B
PPAR alpha
PPAR-beta
PPAR delta
PPAR gamma
Peroxisome Proliferator-Activated Receptors
RNA, Messenger
Tumor Necrosis Factor-alpha
Figure
-
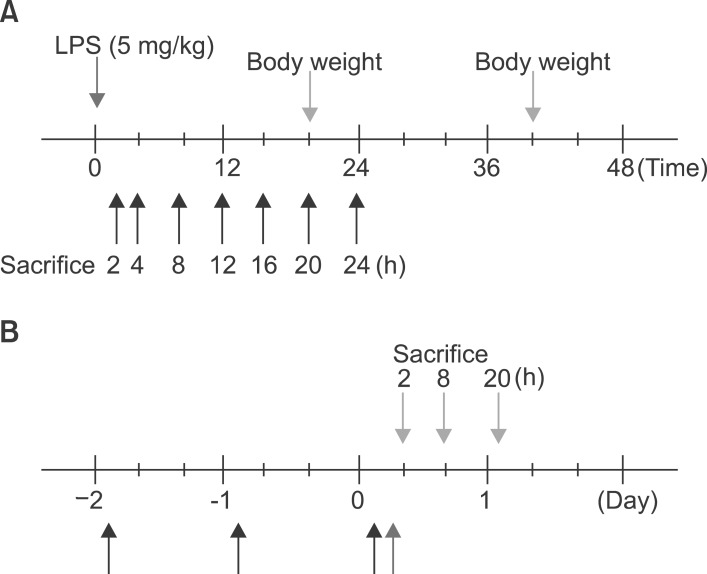
Fig. 1 Schematic representation of in vivo experiment.
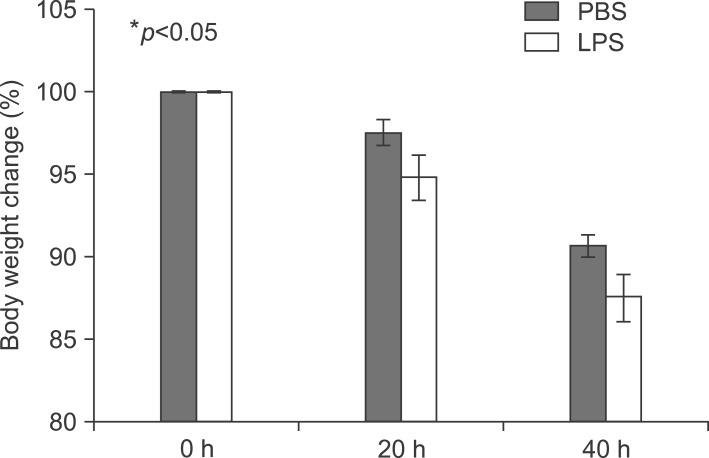
Fig. 2 Body weight change in LPS-injected mice. Balb/c mice were injected intraperitoneally with lipopolysaccharide 5 mg/kg once. Mice were weighed after 20 and 40 h.
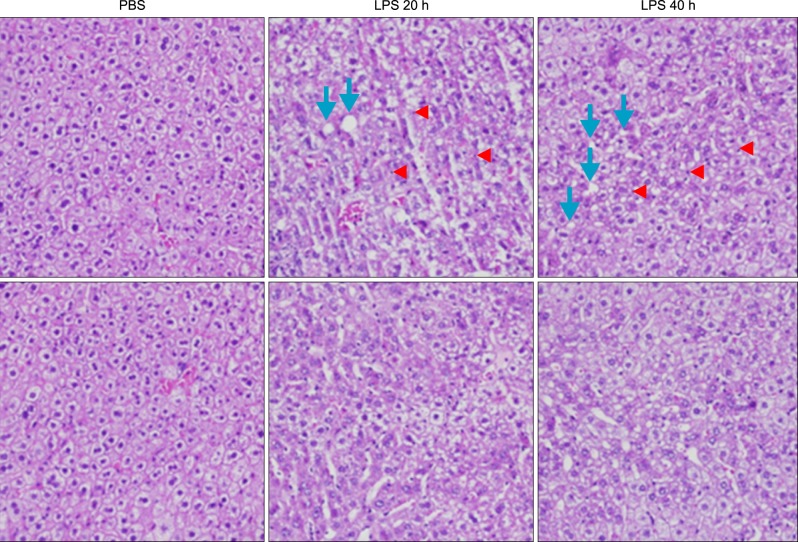
Fig. 3 Histopathological changes of liver sections of mice under LPS treatment. Hematoxylin and eosin stained liver sections from mice liver (original magnification, ×100). Arrowhead indicates leukocytes. Arrow indicates an inflammatory focus.
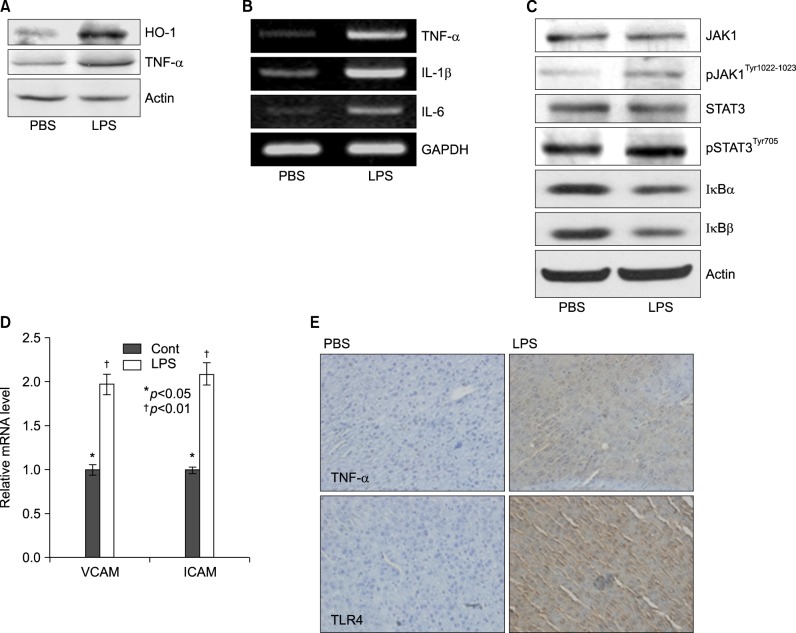
Fig. 4 Lipopolyssacharide-induced inflammation profiles in Balb/c. Mice were injected intraperitoneally with LPS. After 20 h, the liver was collected and analyzed for Western blot, reverse transcriptase PCR, real time PCR, and immunohistochemistry. (A) Induction of pro-inflammatory cytokines, TNF-α, and stress inducible protein, heme oxygenase-1 (HO-1) in LPS-treated mouse. (B) Expression levels of TNF-α, IL-1β, and IL-6 mRNA were determined by RT-PCR in LPS injected mouse liver and observed by agarose-gel electrophoresis under UV illumination staining with ethidium bromide. (C) Liver lysates were prepared. Whole lysates (40 µg of protein) were separated by electrophoresis on SDS 10% polyacrylamide gels and then the protein was transferred onto nitrocellulose membrane. Each membrane was immunoblotted with antibody specific for phosphorylated or tatal JAK and STAT. (D) Levels of VCAM and ICAM mRNA were determined by semi-quantitative real time PCR in LPS injected mouse liver (Cont: *p<0.05, LPS: †p<0.01). (E) Liver from Balb/c mice injected with PBS or LPS were removed and embedded in paraffin, and then 4 µm sections were prepared. For immunohistochemistry studies, an immunohistochemistry kit was used, and all the procedures were performed according to the instructions of the manufacturer. The anti-TNF-α and anti-Tall like receptor (TLR) 4 were used. All results were representative of three separate experiments.
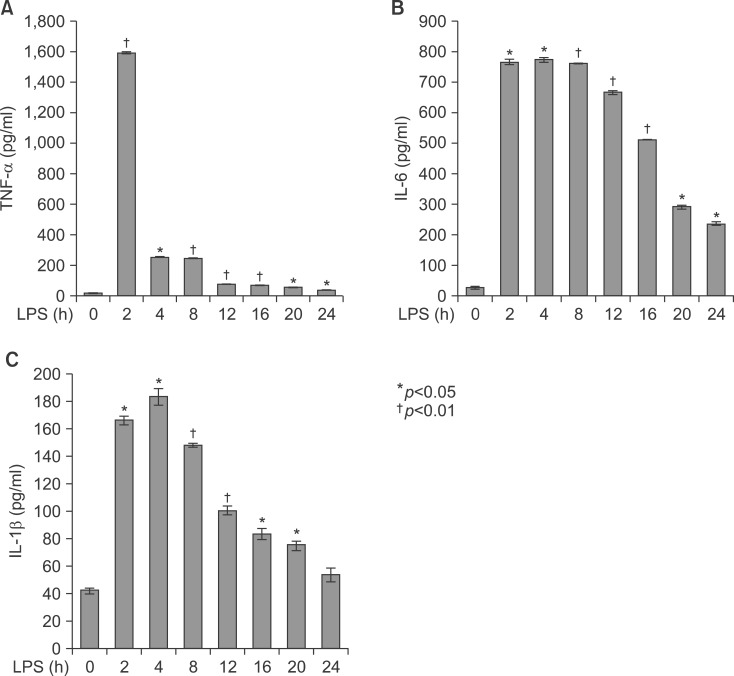
Fig. 5 Secretion of TNF-α, IL-1β, and IL-6 in serum from LPS-injected Balb/c. Secretory protein levels of TNF-α, IL-1β, and IL-6 from the serum were measured by the ELISA method for the indicated times. All results were representative of three separate experiments. (A) injection of LPS resulted in a marked increase of TNF-α which was observed as early as at 2 h and reached a primary peak level with 1,591.15±5.798 pg/ml at 2 h, and thereafter gradually decreased in a time-dependent manner. (B) LPS also increased in secretion of IL-1β, which was already reached a peak level with 183.2604±6.542 pg/ml at 4 h, thereafter sustained its level up to 12 h, and gradually decreased in a time-dependent manner. (C) IL-6 secretion was already reached a peak level with 766.3889±8.214 pg/ml at 2 h, and thereafter gradually decreased in a time-dependent manner.
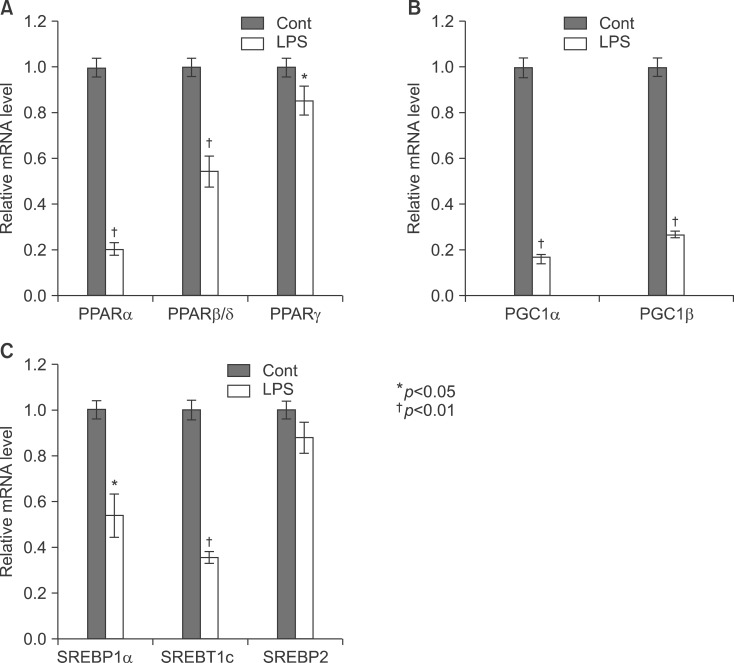
Fig. 6 Expression level of PPARs, PGC1β, ACOX1, SREBP1α, SREBP1c, and SREBP2 mRNA was determined by qPCR (normalized to GAPDH). Balb/c mice were injected with LPS. Total RNA from the liver was isolated. Two micrograms of total RNA was reverse transcribed to cDNA. PPARs (A), PGC1β and ACOX1 (B), SREBP1α, SREBP1c, and SREBP2 (C) mRNA expression was measured by real-time QPCR as described under "Methods". QPCR data were normalized using GAPDH mRNA as the invariant control for all experiments. Data (mean±SD) are expressed as a relative unit.
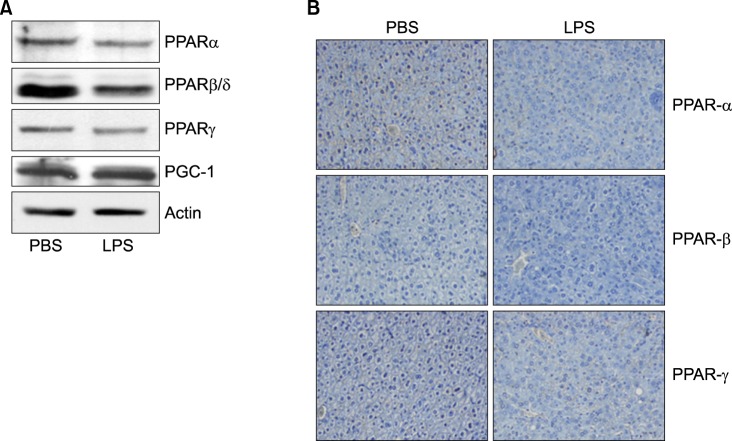
Fig. 7 Expression level of PPARs protein was determined by Western blot and IHC. Mice were injected intraperitoneally with LPS. After 20 h, the liver was collected and analyzed for Western blot (A) and immunohistochemistry (B).
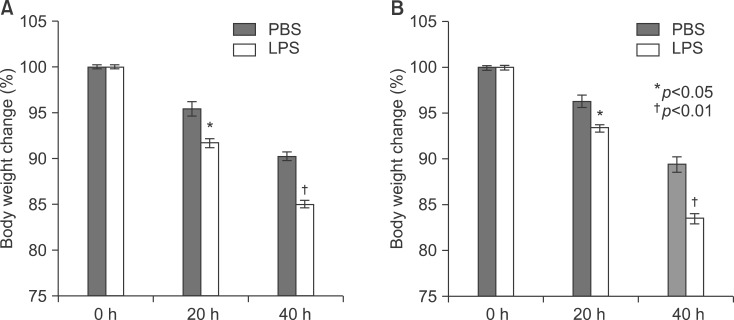
Fig. 8 Body weight change in LPS-injected mice. TNF-α (A) and IL-6 (B) knockout mice were injected intraperitoneally with lipopolysaccharide 5 mg/kg once. Mice were weighed after 20 and 40 h.
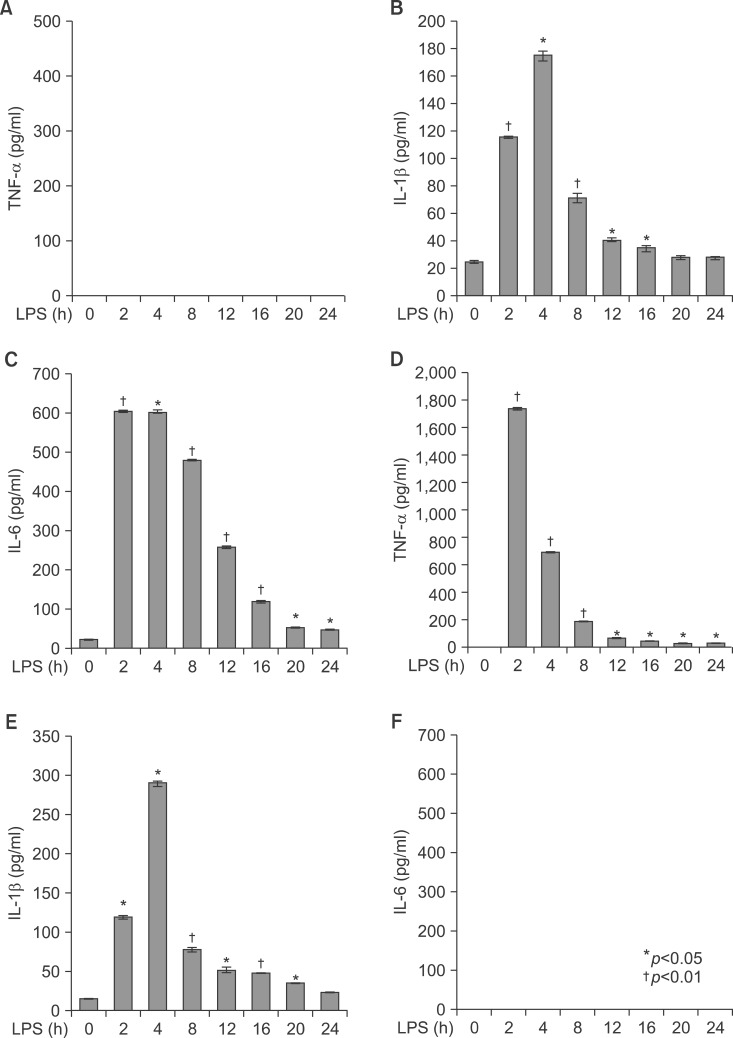
Fig. 9 Secretion of TNF-α, IL-1β, and IL-6 in serum from LPS-injected TNF-α and IL-6 knockout mice. Secretory protein levels of TNF-α, IL-1β, and IL-6 from the serum were measured by the ELISA method for the indicated times. All results were representative of three separate experiments. (A-C) TNF-α knockout mice; (D-F) IL-6 knockout mice.
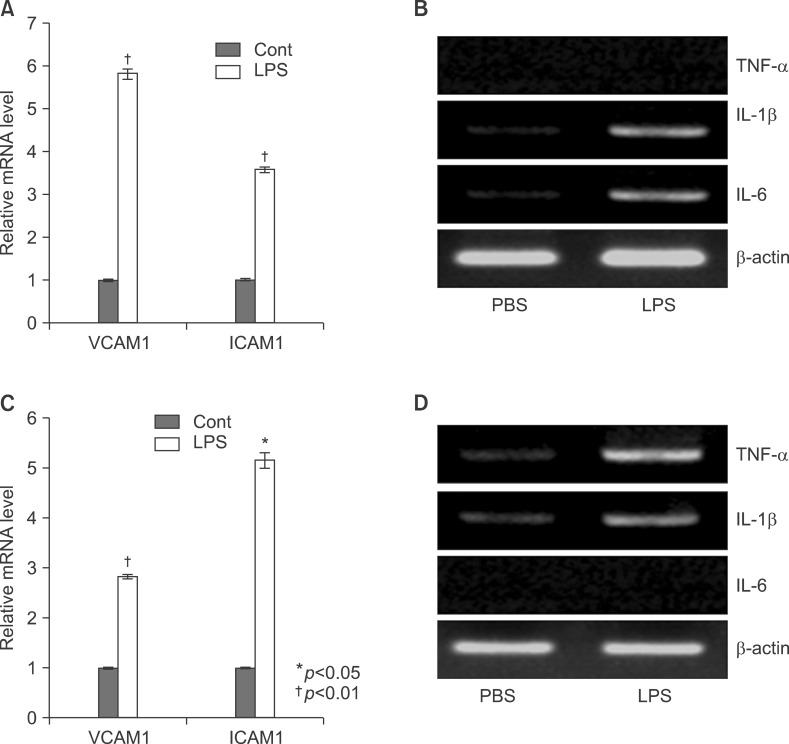
Fig. 10 LPS induced inflammation profiles in TNFa, IL-6 knockout mice. Level of VCAM and ICAM mRNA were determined by semi-quantitative real time PCR in LPS or PBS injected knockout mouse liver (A: TNF-α KO, C: IL-6 KO). Expression levels of TNF-α, IL-1β, and IL-6 mRNA were determined by RT-PCR in LPS injected mouse liver and observed by agarose-gel electrophoresis under UV illumination staining with ethidium bromide (B: TNF-α KO, D: IL-6 KO).
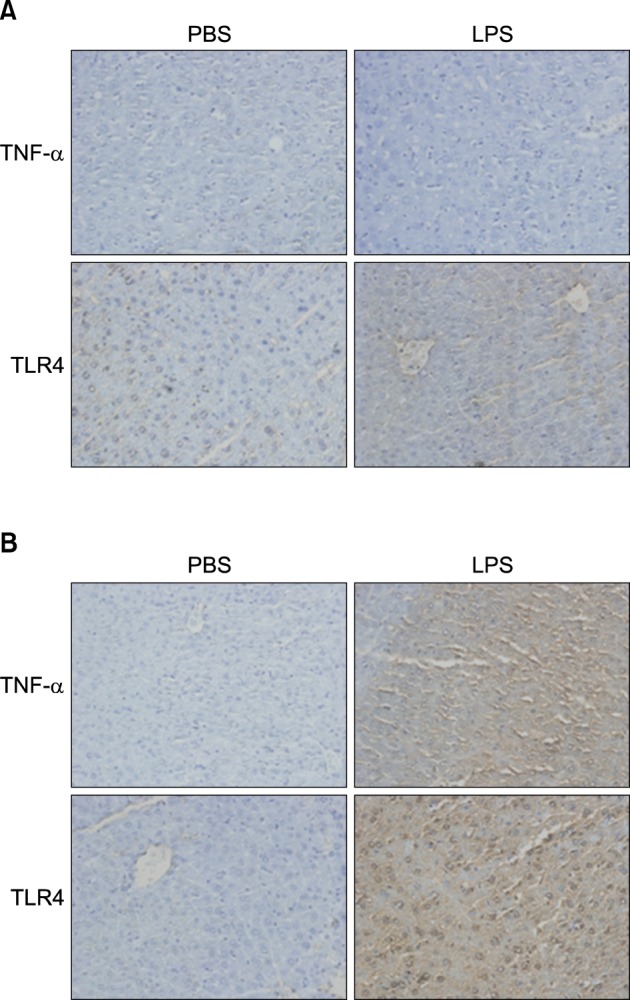
Fig. 11 In vivo effect of LPS on TNF-α and TLR4 expression in the liver of TNF-α (A), IL-6 (B) knockout mice. Liver from knockout mice injected with PBS or LPS were removed and embedded in paraffin, and then 4 µm sections were prepared. For immunohistochemistry studies, an immunohistochemistry kit was used, and all the procedures were performed according to the instructions of the manufacturer. The anti-TNF-α and anti-Tall like receptor (TLR) 4 were used. All results were representative of three separate experiments.
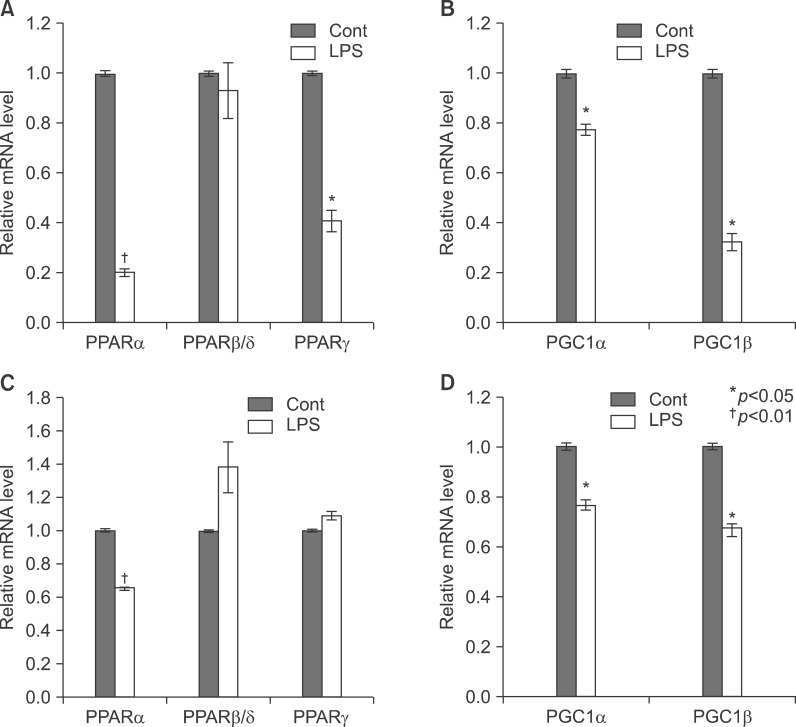
Fig. 12 Expression level of PPARs, PGC1β, ACOX1, SREBP1α, SREBP1c, and SREBP2 mRNA was determined by QPCR (normalized to GAPDH) in the liver of KO mice. TNF-α (A, B) and IL-6 (C, D) KO mice were injected with LPS. Total RNA from liver was isolated. Two micrograms of total RNA was reverse transcribed to cDNA. PPARs (1), PGC1β and ACOX1 (2), SREBP1α, SREBP1c, and SREBP2 (3). mRNA expression was measured by real-time QPCR as described under "Methods." QPCR data were normalized using GAPDH mRNA as the invariant control for all experiments. Data (mean±SD) are expressed as a control level.
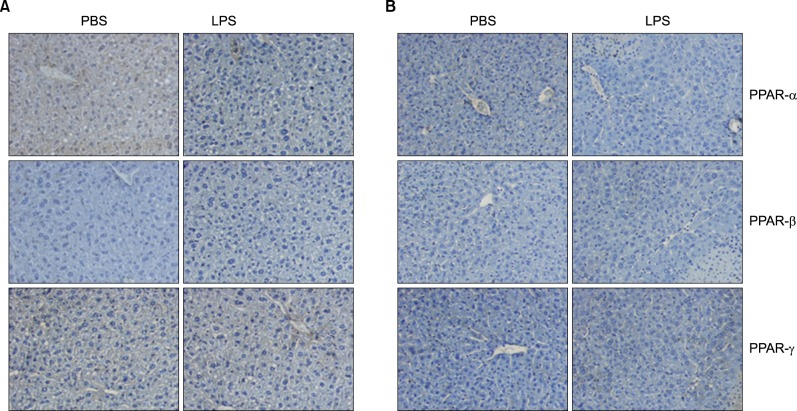
Fig. 13 In vivo effect of LPS on PPARs expression in the liver of TNF-α and IL-6 KO mice. Expression level of PPARs protein was determined by immunohistochemistry. TNF-α (A) and IL-6 KO (B) mice were injected intraperitoneally with LPS. After 20 h, the liver was collected and analyzed for immunohistochemistry.
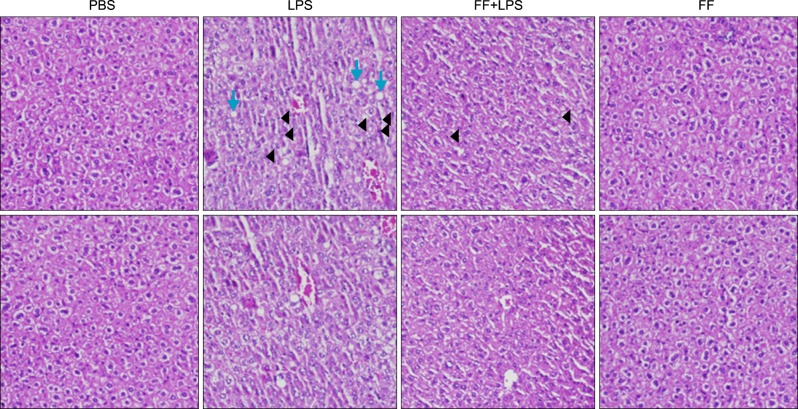
Fig. 14 Effects of fenofibrate on LPS-induced inflammation in Balb/c mice. Hematoxylin and eosin stain (×100) shows intracellular lipid accumulation and inflammation.
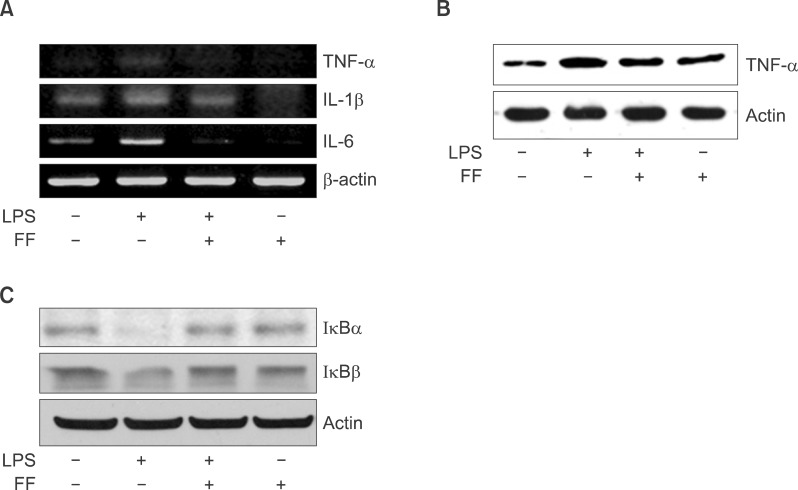
Fig. 15 Effect of fenofibrate on proinflammatory cytokine mRNA and protein level in Balb/c mice. LPS-induced expression levels of mRNA (A) and protein (B) were determined by RT-PCR and Western blot in the presence and absence of fenofibrate as described under "Methods." The amplified gene products were observed by agarose-gel electrophoresis under UV illumination staining with ethidium bromide. TNF-α protein level was detected by Western blot. (C) Western blot analyses of IκBα and IκBβ in liver treated with PPAR-α agonist fenofibrate (100 mg/kg) and followed by LPS treatment (5 mg/kg).
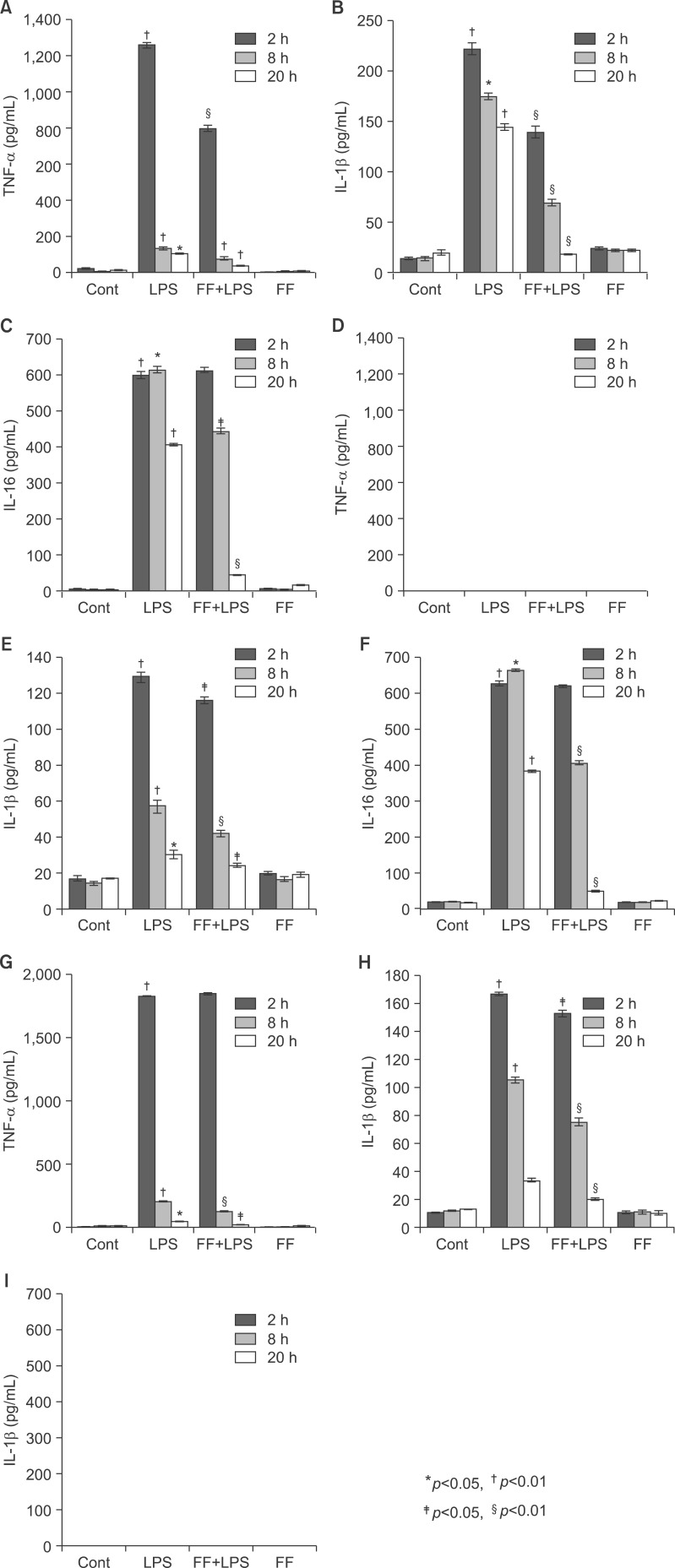
Fig. 16 Effect of fenofibrate on proinflammatory cytokine mRNA levels in serum of Balb/c, TNF-α KO, and IL6 KO mice. All mice were fed fenofibrate (100 mg/kg) or methylcellulose. After LPS injection, the pro-inflammatory cytokines were measured by the ELISA method for the indicated times. All results were representative of three separate experiments. (A-C) Balb/c mouse, (D-F) TNF-α KO mouse; (G-I) IL-6 KO mouse.
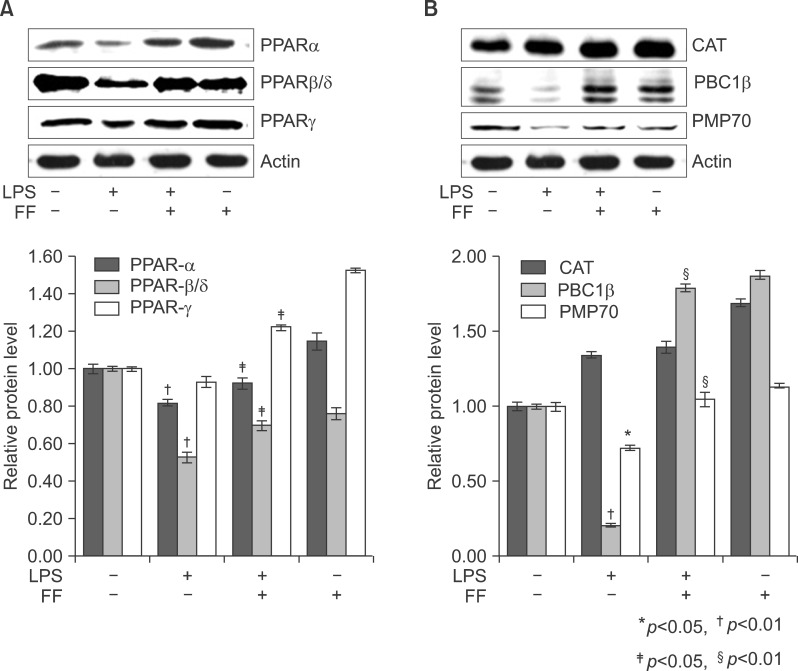
Fig. 17 Effect of fenofibrate on PPARs (A), catalase, PMP70, and PGC1b (B) expression in treated liver tissue of Balb/c. After pretreatment with fenofibrate, LPS was injected intraperitoneally. Whole liver lysates (40 µg of protein) were separated by electrophoresis on SDS 10% polyacrylamide gels and then the protein were transferred onto a nitrocellulose membrane. Each membrane was immunoblotted with antibody specific for PPARs, catalase, PGC1, PMP70, and actin. The results were representative of three separate experiments.
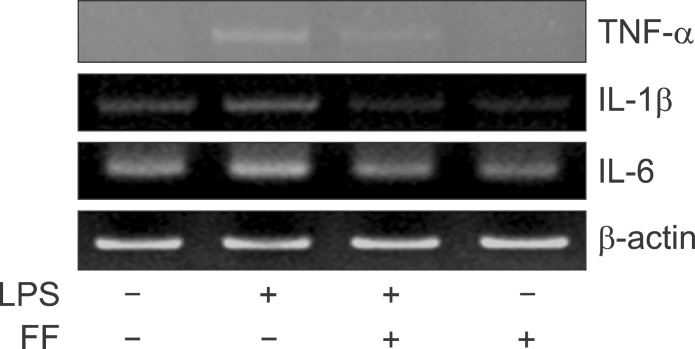
Fig. 18 Effect of fenofibrate on proinflammatory cytokine mRNA level in HepG2. Expression levels of TNF-α, IL-1β, and IL-6 mRNA were determined by RT-PCR. PCR products observed by agarose-gel electrophoresis under UV illumination staining with ethidium bromide (TNF-α KO; IL-6 KO).
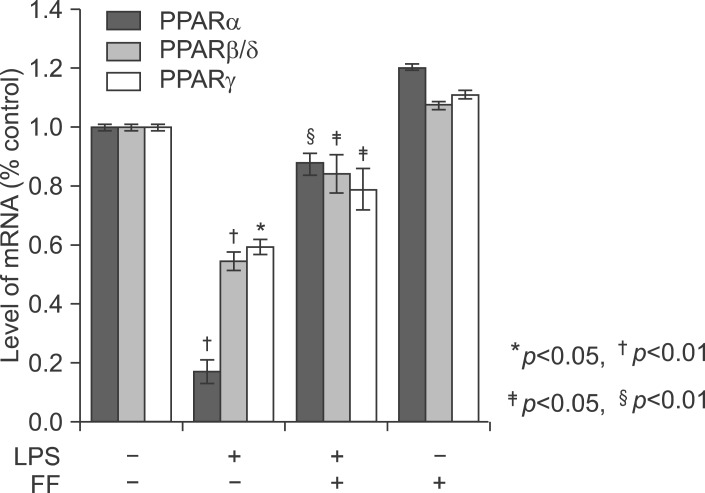
Fig. 19 Effect of fenofibrate on PPARs mRNA level in the liver of Balb/c. Fenofibrate (100 mg/kg) or its vehicle (water containing 0.5% methylcellulose) were administered. After LPS injection, total RNA from the liver was isolated. Two micrograms of total RNA was reverse transcribed to cDNA. PPARs mRNA expression was measured by real-time QPCR as described under "Methods." QPCR data were normalized using GAPDH mRNA as the invariant control for all experiments. Data (mean±SD) are expressed as a relative unit.
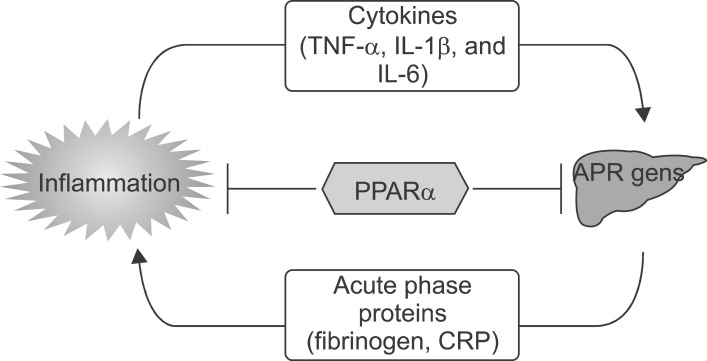
Fig. 20 PPARα is a modulator of the inflammatory response in the liver.
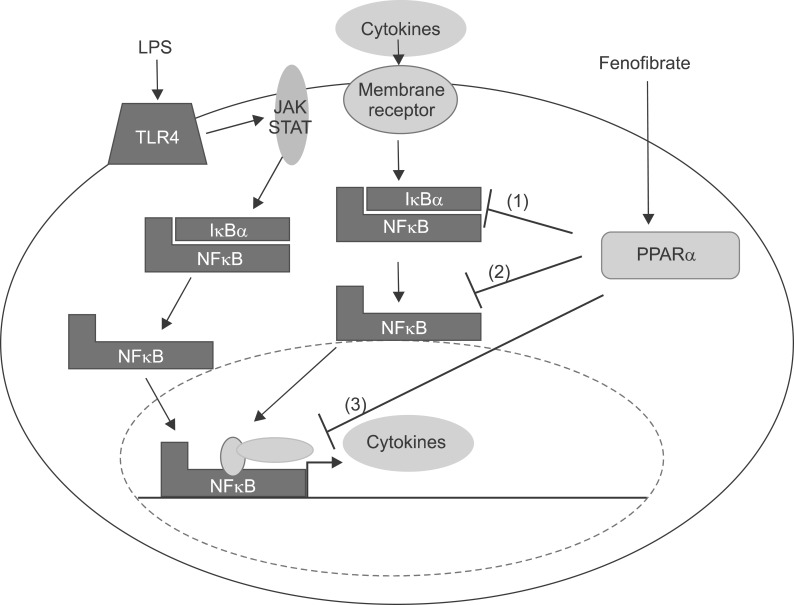
Fig. 21 Cellular mechanism of the LPS in the liver and interaction of PPARα at several levels of the inflammatory signaling pathway. Interaction with the NFκB pathway. LPS increases cytokines including TNF-α, IL-1β, and IL-6 through theNFκB pathway. Fenofibrate, a PPARα agonist, inhibits the NFκB pathway through blocking translocation of NFκB into nucleus. (1) Inhibition of phosphorylation of IκB. (2) Inhibition of nuclear translocation of inflammatory transcription factors. (3) Interference with activation of the transcription initiation complex via cofactor interaction.
Reference
-
1. Streetz KL, Wüstefeld T, Klein C, et al. Mediators of inflammation and acute phase response in the liver. Cell Mol Biol (Noisy-le-grand). 2001; 47:661–673. PMID: 11502073.2. Hoffmeister A, Rothenbacher D, Bäzner U, et al. Role of novel markers of inflammation in patients with stable coronary heart disease. Am J Cardiol. 2001; 87:262–266. PMID: 11165957.
Article3. Ridker PM, Stampfer MJ, Rifai N. Novel risk factors for systemic atherosclerosis: a comparison of C-reactive protein, fibrinogen, homocysteine, lipoprotein(a), and standard cholesterol screening as predictors of peripheral arterial disease. JAMA. 2001; 285:2481–2485. PMID: 11368701.4. Heinrich PC, Behrmann I, Müller-Newen G, et al. Interleukin-6-type cytokine signalling through the gp130/Jak/STAT pathway. Biochem J. 1998; 334:297–314. PMID: 9716487.
Article5. Kim H, Baumann H. Dual signaling role of the protein tyrosine phosphatase SHP-2 in regulating expression of acute-phase plasma proteins by interleukin-6 cytokine receptors in hepatic cells. Mol Cell Biol. 1999; 19:5326–5338. PMID: 10409724.
Article6. Yoo JY, Wang W, Desiderio S, et al. Synergistic activity of STAT3 and c-Jun at a specific array of DNA elements in the alpha 2-macroglobulin promoter. J Biol Chem. 2001; 276:26421–26429. PMID: 11319221.7. Zauberman A, Lapter S, Zipori D. Smad proteins suppress CCAAT/enhancer-binding protein (C/EBP) beta- and STAT3-mediated transcriptional activation of the haptoglobin promoter. J Biol Chem. 2001; 276:24719–24725. PMID: 11331273.8. Issemann I, Green S. Activation of a member of the steroid hormone receptor superfamily by peroxisome proliferators. Nature. 1990; 347:645–650. PMID: 2129546.
Article9. Forman BM, Chen J, Evans RM. Hypolipidemic drugs, polyunsaturated fatty acids, and eicosanoids are ligands for peroxisome proliferator-activated receptors alpha and delta. Proc Natl Acad Sci U S A. 1997; 94:4312–4317. PMID: 9113986.10. Gervois P, Chopin-Delannoy S, Fadel A, et al. Fibrates increase human REV-ERBalpha expression in liver via a novel peroxisome proliferator-activated receptor response element. Mol Endocrinol. 1999; 13:400–409. PMID: 10076997.11. Kersten S, Seydoux J, Peters JM, et al. Peroxisome proliferator-activated receptor alpha mediates the adaptive response to fasting. J Clin Invest. 1999; 103:1489–1498. PMID: 10359558.12. Guerre-Millo M, Gervois P, Raspé E, et al. Peroxisome proliferator-activated receptor alpha activators improve insulin sensitivity and reduce adiposity. J Biol Chem. 2000; 275:16638–16642. PMID: 10828060.13. Gervois P, Torra IP, Fruchart JC, et al. Regulation of lipid and lipoprotein metabolism by PPAR activators. Clin Chem Lab Med. 2000; 38:3–11. PMID: 10774955.
Article14. Gervois P, Kleemann R, Pilon A, et al. Global suppression of IL-6-induced acute phase response gene expression after chronic in vivo treatment with the peroxisome proliferator-activated receptor-alpha activator fenofibrate. J Biol Chem. 2004; 279:16154–16160. PMID: 14764586.15. Stienstra R, Mandard S, Patsouris D, et al. Peroxisome proliferator-activated receptor alpha protects against obesity-induced hepatic inflammation. Endocrinology. 2007; 148:2753–2763. PMID: 17347305.16. He W, Barak Y, Hevener A, et al. Adipose-specific peroxisome proliferator-activated receptor gamma knockout causes insulin resistance in fat and liver but not in muscle. Proc Natl Acad Sci U S A. 2003; 100:15712–15717. PMID: 14660788.17. Marra F, Efsen E, Romanelli RG, et al. Ligands of peroxisome proliferator-activated receptor gamma modulate profibrogenic and proinflammatory actions in hepatic stellate cells. Gastroenterology. 2000; 119:466–478. PMID: 10930382.18. Daynes RA, Jones DC. Emerging roles of PPARs in inflammation and immunity. Nat Rev Immunol. 2002; 2:748–759. PMID: 12360213.
Article19. Planavila A, Rodríguez-calvo R, Jové M, et al. Peroxisome proliferator-activated receptor beta/delta activation inhibits hypertrophy in neonatal rat cardiomyocytes. Cardiovasc Res. 2005; 65:832–841. PMID: 15721863.20. Planavila A, Laguna JC, Vázquez-carrera M. Nuclear factor-kappaB activation leads to down-regulation of fatty acid oxidation during cardiac hypertrophy. J Biol Chem. 2005; 280:17464–17471. PMID: 15728586.21. Xu J, Storer PD, Chavis JA, et al. Agonists for the peroxisome proliferator-activated receptor-alpha and the retinoid X receptor inhibit inflammatory responses of microglia. J Neurosci Res. 2005; 81:403–411. PMID: 15968640.22. Jonkers IJ, Mohrschladt MF, Westendorp RG, et al. Severe hypertriglyceridemia with insulin resistance is associated with systemic inflammation: reversal with bezafibrate therapy in a randomized controlled trial. Am J Med. 2002; 112:275–280. PMID: 11893366.
Article23. Khovidhunkit W, Kim MS, Memon RA, et al. Effects of infection and inflammation on lipid and lipoprotein metabolism: mechanisms and consequences to the host. J Lipid Res. 2004; 45:1169–1196. PMID: 15102878.24. Zhong Z, Wen Z, Darnell JE Jr. Stat3 and Stat4: members of the family of signal transducers and activators of transcription. Proc Natl Acad Sci USA. 1994; 91:4806–4810. PMID: 7545930.
Article25. Devchand PR, Keller H, Peters JM, et al. The PPARalpha-leukotriene B4 pathway to inflammation control. Nature. 1996; 384:39–43. PMID: 8900274.26. Jackson SM, Parhami F, Xi XP, et al. Peroxisome proliferator-activated receptor activators target human endothelial cells to inhibit leukocyte-endothelial cell interaction. Arterioscler Thromb Vasc Biol. 1999; 19:2094–2104. PMID: 10479650.
Article27. Marx N, Mackman N, Schönbeck U, et al. PPARalpha activators inhibit tissue factor expression and activity in human monocytes. Circulation. 2001; 103:213–219. PMID: 11208679.28. Neve BP, Corseaux D, Chinetti G, et al. PPARalpha agonists inhibit tissue factor expression in human monocytes and macrophages. Circulation. 2001; 103:207–212. PMID: 11208678.29. Sirtori CR, Colli S. Influences of lipid-modifying agents on hemostasis. Cardiovasc Drugs Ther. 1993; 7:817–823. PMID: 8110626.
Article30. Gervois P, Vu-dac N, Kleemann R, et al. Negative regulation of human fibrinogen gene expression by peroxisome proliferator-activated receptor alpha agonists via inhibition of CCAAT box/enhancer-binding protein beta. J Biol Chem. 2001; 276:33471–33477. PMID: 11418615.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Regulatory Effects of Fenofibrate with Inflammatory Response and Myocardiac Dysfunction in Lipopolysaccharide-stimulated Heart Tissues
- Protective Effects of Fenofibrate Against the Inflammatory Cytokines in Lipopolysaccharide-Induced Mice Brain Tissues
- Refocusing Peroxisome Proliferator Activated Receptor-alpha: A New Insight for Therapeutic Roles in Diabetes
- Peroxisomal Fitness: A Potential Protective Mechanism of Fenofibrate against High Fat Diet-Induced Non-Alcoholic Fatty Liver Disease in Mice
- Safety and Efficacy of Peroxisome Proliferator-Activated Receptor-alpha Agonist for Treating Cardiovascular Disease
