Selective Mitochondrial Uptake of MKT-077 Can Suppress Medullary Thyroid Carcinoma Cell Survival In Vitro and In Vivo
- Affiliations
-
- 1Department of Biochemistry, Medical College of Wisconsin, Milwaukee, WI, USA. jipark@mcw.edu
- KMID: 2169670
- DOI: http://doi.org/10.3803/EnM.2015.30.4.593
Abstract
- BACKGROUND
Medullary thyroid carcinoma (MTC) is a neuroendocrine tumor mainly caused by mutations in the rearranged during transfection (RET) proto-oncogene. Not all patients with progressive MTC respond to current therapy inhibiting RET, demanding additional therapeutic strategies. We recently demonstrated that disrupting mitochondrial metabolism using a mitochondria-targeted agent or by depleting a mitochondrial chaperone effectively suppressed human MTC cells in culture and in mouse xenografts by inducing apoptosis and RET downregulation. These observations led us to hypothesize that mitochondria are potential therapeutic targets for MTC. This study further tests this hypothesis using1-ethyl-2-[[3-ethyl-5-(3-methylbenzothiazolin-2-yliden)]-4-oxothiazolidin-2-ylidenemethyl] pyridinium chloride (MKT-077), a water-soluble rhodocyanine dye analogue, which can selectively accumulate in mitochondria.
METHODS
The effects of MKT-077 on cell proliferation, survival, expression of RET and tumor protein 53 (TP53), and mitochondrial activity were determined in the human MTC lines in culture and in mouse xenografts.
RESULTS
MKT-077 induced cell cycle arrest in TT and MZ-CRC-1. Intriguingly, MKT-077 also induced RET downregulation and strong cell death responses in TT cells, but not in MZ-CRC-1 cells. This discrepancy was mainly due to the difference between the capacities of these cell lines to retain MKT-077 in mitochondria. The cytotoxicity of MKT-077 in TT cells was mainly attributed to oxidative stress while being independent of TP53. MKT-077 also effectively suppressed tumor growth of TT xenografts.
CONCLUSION
MKT-077 can suppress cell survival of certain MTC subtypes by accumulating in mitochondria and interfering with mitochondrial activity although it can also suppress cell proliferation via other mechanisms. These results consistently support the hypothesis that mitochondrial targeting has therapeutic potential for MTC.
Keyword
MeSH Terms
Figure
-
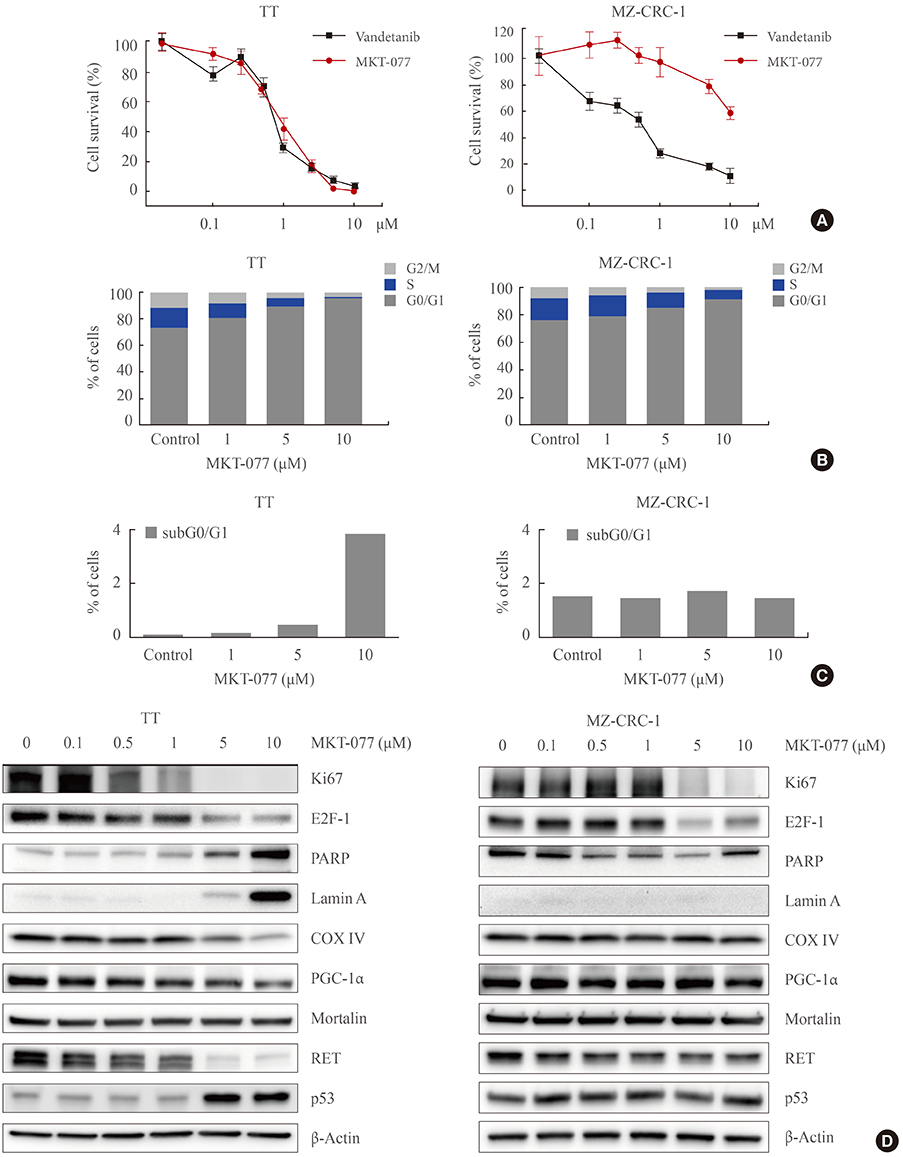
Fig. 1 1-Ethyl-2-[[3-ethyl-5-(3-methylbenzothiazolin-2-yliden)]-4-oxothiazolidin-2-ylidenemethyl] pyridinium chloride (MKT-077) induces growth inhibitory effects in medullary thyroid carcinoma (MTC) cell culture. (A) TT and MZ-CRC-1 cells in 24-well plates were treated with serially increasing doses of MKT-077 and vandetanib for 48 hours. Cells were then allowed to recover in drug-free fresh medium for 48 hours, prior to measurement of cell viability by MTT assay. Data (mean±SD, n=6) are expressed as the percentage of vehicle-treated control. (B, C) TT and MZ-CRC-1 cells were treated with MKT-077 at indicated doses for 48 hours prior to cell cycle analysis using Hoechst 33342. Control cells were treated with equal volume of dimethyl sulfoxide. (B) Data are expressed as the percentage of living cells in each cell cycle phase. (C) Number of apoptotic cells was estimated by counting cells in the subG0/G1 phase. (D) Total lysates of TT and MZ-CRC-1 cells treated with different doses of MKT-077 for 48 hours were analyzed by Western blotting for indicated proteins. β-Actin is the control for equal loading. Ki67, MKI67-encoded protein; E2F-1, E2 promoter binding factor 1; PARP, poly(ADP-ribose) polymerase; COX IV, cytochrome c oxidase; PGC-1α, peroxisome proliferator-activated receptor gamma coactivator 1α; RET, rearranged during transfection.

Fig. 2 Differential mitochondrial accumulation of 1-ethyl-2-[[3-ethyl-5-(3-methylbenzothiazolin-2-yliden)]-4-oxothiazolidin-2-ylidenemethyl] pyridinium chloride (MKT-077) in medullary thyroid carcinoma (MTC) cells. (A) TT and MZ-CRC-1 cells were treated with 1 µM MKT-077 and 100 nM MitoTracker Green FM (Thermo Fisher Scientific) for 30 minutes. Pictures were then taken under a fluorescent microscope (×400). (B, C) TT and MZ-CRC-1 cells, treated with increasing doses of MKT-077 for 1 hour, were analyzed by flow cytometry to measure red fluorescence (PE channel, 575 nm) (B). (C) Data of 20,000 cells per dose were normalized to dimethyl sulfoxide-treated control and presented as mean±SD. (D) TT and MZ-CRC-1 cells were stained with 5 nM TMRE for 15 minutes and analyzed by flow cytometry to measure red fluorescence (PE channel, 575 nm). Unstained cells were used as negative controls. (E) Total lysates of TT and MZ-CRC-1 cells were analyzed by Western blotting for indicated proteins. GAPDH is the control for equal loading. TMRE, tetra-methyl-rhodamine ethyl ester perchlorate; RET, rearranged during transfection; PGC-1α, peroxisome proliferator-activated receptor gamma coactivator 1α; COX IV, cytochrome c oxidase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.

Fig. 3 1-Ethyl-2-[[3-ethyl-5-(3-methylbenzothiazolin-2-yliden)]-4-oxothiazolidin-2-ylidenemethyl] pyridinium chloride (MKT-077) effects are mediated independently of TP53. (A) Western blot analysis of total lysates. TT cells were infected for 5 days with lentiviral pLKO.1 expressing TP53-targeting shRNA (shp53) or the control virus pLKO.1 prior to the treatment with indicated doses of MKT-077 for 48 hours. (B) TT cells infected with lentiviral shp53 or the control pLKO.1 in 24-well plates were treated with increasing doses of MKT-077 for 48 hours. Cells were then allowed to recover in drug-free fresh medium for 48 hours prior to the measurement of cell viability by MTT assay. Data (mean±SD, n=4) are expressed as the percentage of vehicle-treated control. Ki67, MKI67-encoded protein; E2F-1, E2 promoter binding factor 1; RET, rearranged during transfection; PARP, poly(ADP-ribose) polymerase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; NS, not significant.

Fig. 4 The reactive oxygen species (ROS) scavenger, N-acetyl-cysteine (NAC), can attenuate 1-ethyl-2-[[3-ethyl-5-(3-methylbenzothiazolin-2-yliden)]-4-oxothiazolidin-2-ylidenemethyl] pyridinium chloride (MKT-077)-induced ROS production and cell death, but not RET down-regulation. (A) TT cells treated with MKT-077 for 24 hours were treated with 1 µM 5-(and-6)-carboxy-2',7'-dichlorodihydrofluorescein diacetate (carboxy-H2DCFDA) for 1 hour and then incubated for 2 hours in a dye-free culture medium. Cells were harvested and analyzed by flow cytometry to measure green fluorescence (fluorescein isothiocyanate [FITC] channel, 525 nm). (B) Cells were pretreated with 5 mM NAC for 2 hours, and then incubated with indicated doses of MKT-077 for 24 hours before measurement of carboxy-H2DCFDA fluorescence by flow cytometry. (C) Cells were treated with indicated doses of MKT-077 in the presence of 5 mM NAC in the medium containing 2% fetal bovine serum (FBS) for 24 hours. Total cell lysates were analyzed by Western blotting for indicated proteins. GAPDH is the control for equal loading. PARP, poly(ADP-ribose) polymerase; RET, rearranged during transfection; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.

Fig. 5 1-Ethyl-2-[[3-ethyl-5-(3-methylbenzothiazolin-2-yliden)]-4-oxothiazolidin-2-ylidenemethyl] pyridinium chloride (MKT-077) suppresses TT xenografts in athymic mice. (A) Athymic mice bearing TT xenografts were treated with MKT-077 (10 mg/kg/dose). Drug was administered intraperitoneal every two days beginning from day 22 after tumor implantation. The control group was treated with the vehicle only (METHODS). Changes in tumor sizes were determined at indicated time-points. (B) Weights of tumors collected at the end of the treatment. Data are mean±SE (n=8). aP<0.05; bP<0.001.
Reference
-
1. Tuttle RM, Ball DW, Byrd D, Daniels GH, Dilawari RA, Doherty GM, et al. Medullary carcinoma. J Natl Compr Canc Netw. 2010; 8:512–530.2. Agrawal N, Jiao Y, Sausen M, Leary R, Bettegowda C, Roberts NJ, et al. Exomic sequencing of medullary thyroid cancer reveals dominant and mutually exclusive oncogenic mutations in RET and RAS. J Clin Endocrinol Metab. 2013; 98:E364–E369.3. Boichard A, Croux L, Al Ghuzlan A, Broutin S, Dupuy C, Leboulleux S, et al. Somatic RAS mutations occur in a large proportion of sporadic RET-negative medullary thyroid carcinomas and extend to a previously unidentified exon. J Clin Endocrinol Metab. 2012; 97:E2031–E2035.4. Ciampi R, Mian C, Fugazzola L, Cosci B, Romei C, Barollo S, et al. Evidence of a low prevalence of RAS mutations in a large medullary thyroid cancer series. Thyroid. 2013; 23:50–57.5. Ichihara M, Murakumo Y, Takahashi M. RET and neuroendocrine tumors. Cancer Lett. 2004; 204:197–211.6. Degrauwe N, Sosa JA, Roman S, Deshpande HA. Vandetanib for the treatment of metastatic medullary thyroid cancer. Clin Med Insights Oncol. 2012; 6:243–252.7. Nagilla M, Brown RL, Cohen EE. Cabozantinib for the treatment of advanced medullary thyroid cancer. Adv Ther. 2012; 29:925–934.8. Wells SA Jr, Robinson BG, Gagel RF, Dralle H, Fagin JA, Santoro M, et al. Vandetanib in patients with locally advanced or metastatic medullary thyroid cancer: a randomized, double-blind phase III trial. J Clin Oncol. 2012; 30:134–141.9. Ward PS, Thompson CB. Metabolic reprogramming: a cancer hallmark even warburg did not anticipate. Cancer Cell. 2012; 21:297–308.10. Sabharwal SS, Schumacker PT. Mitochondrial ROS in cancer: initiators, amplifiers or an Achilles' heel? Nat Rev Cancer. 2014; 14:709–721.11. Don AS, Hogg PJ. Mitochondria as cancer drug targets. Trends Mol Med. 2004; 10:372–378.12. Weinberg SE, Chandel NS. Targeting mitochondria metabolism for cancer therapy. Nat Chem Biol. 2015; 11:9–15.13. Starenki D, Park JI. Mitochondria-targeted nitroxide, Mito-CP, suppresses medullary thyroid carcinoma cell survival in vitro and in vivo. J Clin Endocrinol Metab. 2013; 98:1529–1540.14. Starenki D, Hong SK, Lloyd RV, Park JI. Mortalin (GRP75/HSPA9) upregulation promotes survival and proliferation of medullary thyroid carcinoma cells. Oncogene. 2015; 34:4624–4634.15. Koya K, Li Y, Wang H, Ukai T, Tatsuta N, Kawakami M, et al. MKT-077, a novel rhodacyanine dye in clinical trials, exhibits anticarcinoma activity in preclinical studies based on selective mitochondrial accumulation. Cancer Res. 1996; 56:538–543.16. Chiba Y, Kubota T, Watanabe M, Matsuzaki SW, Otani Y, Teramoto T, et al. MKT-077, localized lipophilic cation: antitumor activity against human tumor xenografts serially transplanted into nude mice. Anticancer Res. 1998; 18(2A):1047–1052.17. Propper DJ, Braybrooke JP, Taylor DJ, Lodi R, Styles P, Cramer JA, et al. Phase I trial of the selective mitochondrial toxin MKT077 in chemo-resistant solid tumours. Ann Oncol. 1999; 10:923–927.18. Rousaki A, Miyata Y, Jinwal UK, Dickey CA, Gestwicki JE, Zuiderweg ER. Allosteric drugs: the interaction of antitumor compound MKT-077 with human Hsp70 chaperones. J Mol Biol. 2011; 411:614–632.19. Arthan D, Hong SK, Park JI. Leukemia inhibitory factor can mediate Ras/Raf/MEK/ERK-induced growth inhibitory signaling in medullary thyroid cancer cells. Cancer Lett. 2010; 297:31–41.20. Park JI, Strock CJ, Ball DW, Nelkin BD. The Ras/Raf/MEK/extracellular signal-regulated kinase pathway induces autocrine-paracrine growth inhibition via the leukemia inhibitory factor/JAK/STAT pathway. Mol Cell Biol. 2003; 23:543–554.21. Park JI, Strock CJ, Ball DW, Nelkin BD. Interleukin-1beta can mediate growth arrest and differentiation via the leukemia inhibitory factor/JAK/STAT pathway in medullary thyroid carcinoma cells. Cytokine. 2005; 29:125–134.22. Hong SK, Yoon S, Moelling C, Arthan D, Park JI. Noncatalytic function of ERK1/2 can promote Raf/MEK/ERK-mediated growth arrest signaling. J Biol Chem. 2009; 284:33006–33018.23. Wu PK, Hong SK, Veeranki S, Karkhanis M, Starenki D, Plaza JA, et al. A mortalin/HSPA9-mediated switch in tumor-suppressive signaling of Raf/MEK/extracellular signal-regulated kinase. Mol Cell Biol. 2013; 33:4051–4067.24. Rubinson DA, Dillon CP, Kwiatkowski AV, Sievers C, Yang L, Kopinja J, et al. A lentivirus-based system to functionally silence genes in primary mammalian cells, stem cells and transgenic mice by RNA interference. Nat Genet. 2003; 33:401–406.25. de Groot JW, Plaza Menacho I, Schepers H, Drenth-Diephuis LJ, Osinga J, Plukker JT, et al. Cellular effects of imatinib on medullary thyroid cancer cells harboring multiple endocrine neoplasia type 2A and 2B associated RET mutations. Surgery. 2006; 139:806–814.26. Morisi R, Celano M, Tosi E, Schenone S, Navarra M, Ferretti E, et al. Growth inhibition of medullary thyroid carcinoma cells by pyrazolo-pyrimidine derivates. J Endocrinol Invest. 2007; 30:RC31–RC34.27. Plaza-Menacho I, Mologni L, Sala E, Gambacorti-Passerini C, Magee AI, Links TP, et al. Sorafenib functions to potently suppress RET tyrosine kinase activity by direct enzymatic inhibition and promoting RET lysosomal degradation independent of proteasomal targeting. J Biol Chem. 2007; 282:29230–29240.28. Rosen A, Casciola-Rosen L. Macromolecular substrates for the ICE-like proteases during apoptosis. J Cell Biochem. 1997; 64:50–54.29. Modica-Napolitano JS, Aprille JR. Delocalized lipophilic cations selectively target the mitochondria of carcinoma cells. Adv Drug Deliv Rev. 2001; 49:63–70.30. Chunta JL, Vistisen KS, Yazdi Z, Braun RD. Uptake rate of cationic mitochondrial inhibitor MKT-077 determines cellular oxygen consumption change in carcinoma cells. PLoS One. 2012; 7:e37471.31. Perry SW, Norman JP, Barbieri J, Brown EB, Gelbard HA. Mitochondrial membrane potential probes and the proton gradient: a practical usage guide. Biotechniques. 2011; 50:98–115.32. Wadhwa R, Sugihara T, Yoshida A, Nomura H, Reddel RR, Simpson R, et al. Selective toxicity of MKT-077 to cancer cells is mediated by its binding to the hsp70 family protein mot-2 and reactivation of p53 function. Cancer Res. 2000; 60:6818–6821.33. Lu WJ, Lee NP, Kaul SC, Lan F, Poon RT, Wadhwa R, et al. Mortalin-p53 interaction in cancer cells is stress dependent and constitutes a selective target for cancer therapy. Cell Death Differ. 2011; 18:1046–1056.34. Green DR, Reed JC. Mitochondria and apoptosis. Science. 1998; 281:1309–1312.35. Halliwell B. Oxidative stress and cancer: have we moved forward? Biochem J. 2007; 401:1–11.36. Kalyanaraman B. Oxidative chemistry of fluorescent dyes: implications in the detection of reactive oxygen and nitrogen species. Biochem Soc Trans. 2011; 39:1221–1225.37. Klein P, Muller-Rischart AK, Motori E, Schonbauer C, Schnorrer F, Winklhofer KF, et al. Ret rescues mitochondrial morphology and muscle degeneration of Drosophila Pink1 mutants. EMBO J. 2014; 33:341–355.38. Kaul SC, Reddel RR, Mitsui Y, Wadhwa R. An N-terminal region of mot-2 binds to p53 in vitro. Neoplasia. 2001; 3:110–114.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Concurrent Medullay and Papillary Carcinoma of the Thyroid
- Medullary Thyroid Carcinoma with Normal Calcitonin Level
- Medullary Carcinoma of Thyroid Gland with Co-existing Papillary Carcinoma
- Concurrent Papillary and Medullary Carcinoma of the Thyroid Gland
- Concurrent Medullary Thyroid Carcinoma and Primary Thyroid Lymphoma (Diffuse Large B Cell Lymphoma): the First Case Report
