Enhanced Allergic Inflammation of Der p 2 Affected by Polymorphisms of MD-2 Promoter
- Affiliations
-
- 1Center for Translational Medicine, Department of Medical Research, Taichung Veterans General Hospital, Taichung, Taiwan.
- 2Department of BioIndustry Technology, Da Yeh University, Changhua, Taiwan.
- 3Department of Medical Technology, Jen Ten College of Medicine, Nursing and Management, Miaoli, Taiwan.
- 4Division of Allergy, Immunology & Rheumatology, Department of Internal Medicine, Taichung Veterans General Hospital, Taichung, Taiwan. jawji@vghtc.gov.tw
- 5College of Life Sciences, National Chung Hsing University, Taichung, Taiwan.
- 6Institute of Clinical Medicine, National Yang Ming University, Taipei, Taiwan.
- KMID: 2147959
- DOI: http://doi.org/10.4168/aair.2015.7.5.497
Abstract
- PURPOSE
Myeloid differentiation-2 (MD-2) has been associated with endotoxin and inflammatory disorders because it can recognize lipopolysaccharide (LPS) binding and attenuate Toll-like receptor 4 (TLR4)-mediated signaling. However, its role in allergic inflammation has yet to be clarified. We examined whether single nucleotide polymorphisms (SNPs) in MD-2 promoter can affect MD-2 expression and aimed to clarify the relationship between Der p 2 allergy and SNPs of MD-2 promoter.
METHODS
The function of SNPs of MD-2 promoter and the effects of cytokines and immunoglobulin on the secretion and mRNA expression were investigated in 73 allergic subjects with different MD-2 gene promoter variants. Peripheral blood mononuclear cells were cultured with or without LPS in the presence of Dermatophagoides pteronyssinus group 2 allergen (Der p 2), followed by mRNA extraction and cytokine expression analysis. The culture supernatants were collected for cytokine measurement.
RESULTS
Patients with the MD-2 promoter SNPs (rs1809441/rs1809442) had increased mRNA expressions of MD-2, epsilon heavy chain of IgE (Cepsilon), and interleukin (IL)-8; however, only MD-2 and IL-8 were further up-regulated after Der p 2 stimulation. Patients with SNPs of MD-2 promoter tended to have high levels of IL-1beta, IL-6, IL-8, IL-10, and tumor necrosis factor (TNF)-alpha after Der p 2 and LPS stimulation. Increased secretions of IL-6, IL-8, and IL-10 were found to be up-regulated by Der p 2 stimulation, and an increased secretion of IFN-gamma and decreased secretion of IL-4 were noted after LPS stimulation.
CONCLUSIONS
The high levels of proinflammatory cytokines secreted by Der p 2 were predetermined by MD-2 promoter SNPs (rs1809441/rs1809442). Through cytokine secretion by Der p 2 and LPS, these SNPs may serve as an indicator of the pathological phenotype of Der p 2-induced allergic inflammation.
Keyword
MeSH Terms
-
Cytokines
Dermatophagoides pteronyssinus
Humans
Hypersensitivity
Immunoglobulin E
Immunoglobulins
Inflammation*
Interleukin-10
Interleukin-4
Interleukin-6
Interleukin-8
Interleukins
Phenotype
Polymorphism, Single Nucleotide
RNA, Messenger
Toll-Like Receptor 4
Tumor Necrosis Factor-alpha
Cytokines
Immunoglobulin E
Immunoglobulins
Interleukin-10
Interleukin-4
Interleukin-6
Interleukin-8
Interleukins
RNA, Messenger
Toll-Like Receptor 4
Tumor Necrosis Factor-alpha
Figure
-
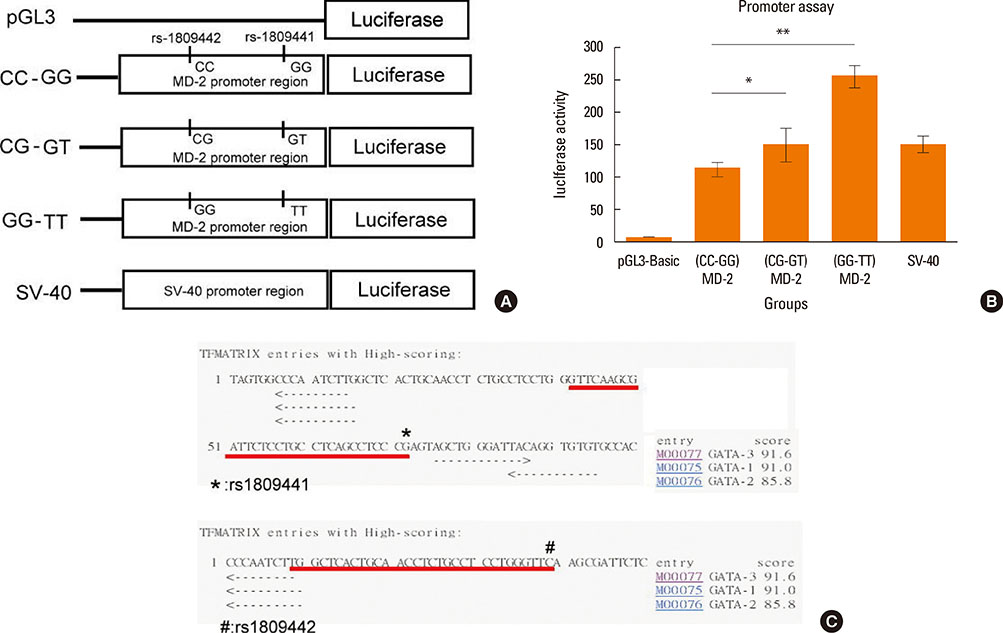
Fig. 1 Effects of MD-2 promoter SNPs on transcription activity. (A) The promoter activity was investigated using a reporter gene assay system. A total of 5 plasmid constructs were prepared by inserting different genotypes of the MD-2 gene promoter region into a pGL3-Basic vector, which contained a promoterless (pGL3-basic), or genotype of CC (rs1809442) GG (rs1809441), or a single mutant genotype of CG and GT, or a combined mutant genotype of GG and TT, or a positive control of SV-40 promoter. (B) Relative luciferase activity (RLA) was assayed in epithelium cells (BEAS-2B) transfected with different plasmid constructs without Der p 2 stimulation. Luciferase activity was normalized for transfection efficiency using a control plasmid (pGL3-Basic). Results are expressed as the fold increase in RLA of the MD-2 promoter construct vector as compared to pGL3-Basic. The values of luciferase activity are expressed as means±SD of the results from each group (n=5). *P<0.05 compared to the MD-2 promoter genotype of rs1809441-G/rs1809442-C constructs; **P<0.01 compared to the MD-2 promoter genotype of rs1809441-G/rs1809442-C constructs. (C) The scoring scheme was performed by simple routine searches highly correlated sequence fragments versus TFMATRIX transcription factor binding site profile database. Score=100.0* ('weight sum' - min)/( max-min); *, sequence position of SNP rs1809441; #, sequence position of SNP rs1809442.
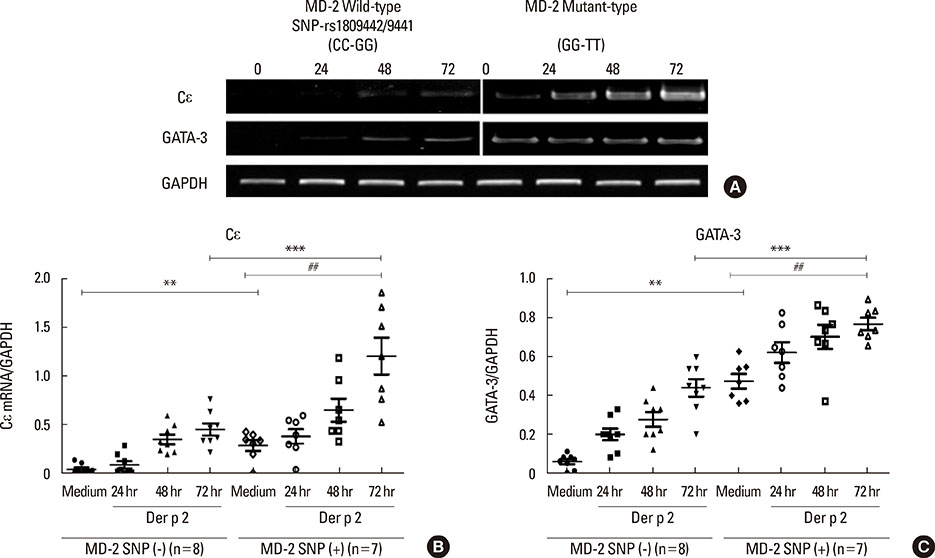
Fig. 2 The mRNA expressions of ε heavy chain of IgE (Cε) and GATA-3 in PBMCs from subjects with the SNP rs1809441/42 mutant and wild genotypes were evaluated using reverse-transcriptase polymerase chain reaction (RT-PCR). (A) The 0, 24, 48, and 72 represent the time points (hours) after allergen rDer p 2 (10 µg/mL) challenge. The GAPDH expression acted as an internal-control. (B) The Cε expression was evaluated using RT-PCR. Data are expressed as mean of each group. Medium, detected without allergen challenge; Der p 2, detected after rDer p 2 (10 µg/mL) challenge. (C) The expression of the transcription factor GATA-3 was evaluated using RT-PCR; **P<0.05 compared between the MD-2 promoter SNPs (+) and SNPs (-) under medium only; ***P<0.01 compared between MD-2 promoter SNPs (+) and SNPs (-) under Der p 2 (10 µg/mL) challenge. ##P<0.05 compared between medium and rDer p 2 (10 µg/mL) challeges in the group of MD-2 promoter SNPs (+).
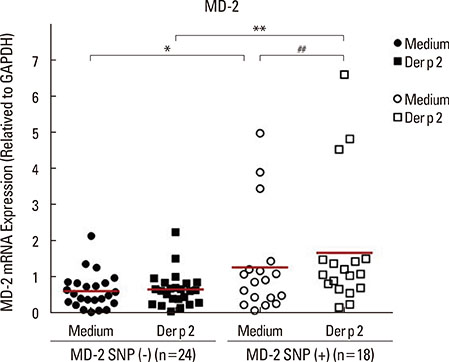
Fig. 3 The SNPs of MD-2 gene promoter affect the MD-2 mRNA expression in PBMC. The MD-2 mRNA expression was evaluated using RT-PCR. Data are expressed as mean of each group. Medium, detected without allergen challenge; Der p 2, detected after the rDer p 2 (10 µg/mL) challenge; *P<0.05 compared between the MD-2 promoter SNPs (+) and SNPs (-) without treatment; **P<0.05 compared between the MD-2 promoter SNPs (+) and SNPs (-) under rDer p 2 challenge; ##P<0.05 compared between medium and rDer p 2 (10 µg/mL) challenge in the group of MD-2 promoter SNPs (+).
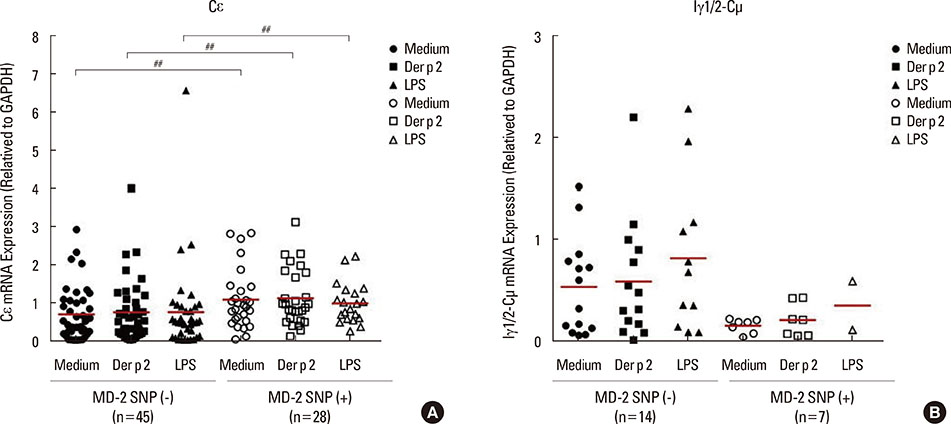
Fig. 4 The SNPs of the MD-2 promoter affect Cε and Iγ1/2-Cµ mRNA expressions in PBMCs. The expressions of ε heavy chain of IgE (Cε) and Iγ1/2-Cµ mRNA were evaluated using RT-PCR. Data are expressed as mean of each group. Medium, detected without treatment; Der p 2, detected after the rDer p 2 (10 µg/mL) challenge; LPS, detected after LPS (100 ng/mL) treatment. ##P<0.05 compared between the MD-2 promoter SNPs (+) and SNPs (-) under different treatments.
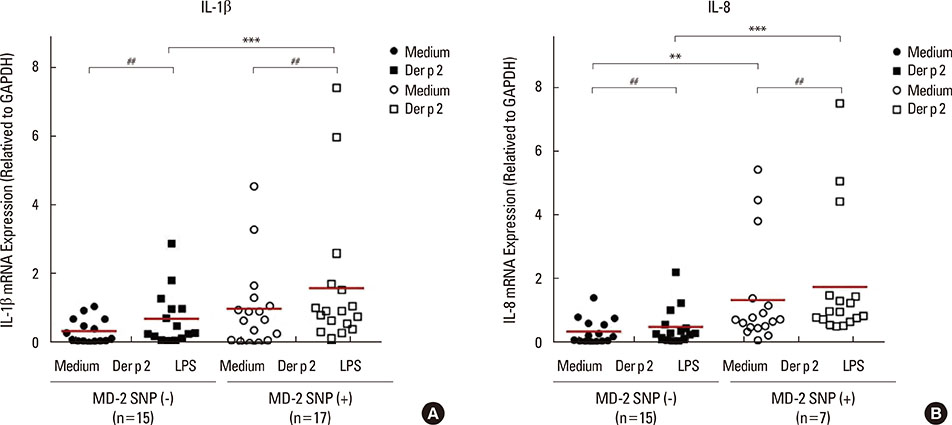
Fig. 5 The SNPs of MD-2 promoter affect the mRNA expressions of IL-1β and IL-8 in PBMCs. The mRNA expressions of IL-1β and IL-8 were evaluated using RT-PCR. Data are expressed as mean of each group. Medium, detected without treatment; Der p 2, detected after the rDer p 2 (10 µg/mL) challenge; ##P<0.05 compared between medium and rDer p 2 challenge; **P<0.05 compared between the MD-2 promoter SNPs (+) and SNPs (-) under medium only; ***P<0.01 compared between MD-2 promoter SNPs (+) and SNPs (-) under Der p 2 (10 µg/mL) challenge.
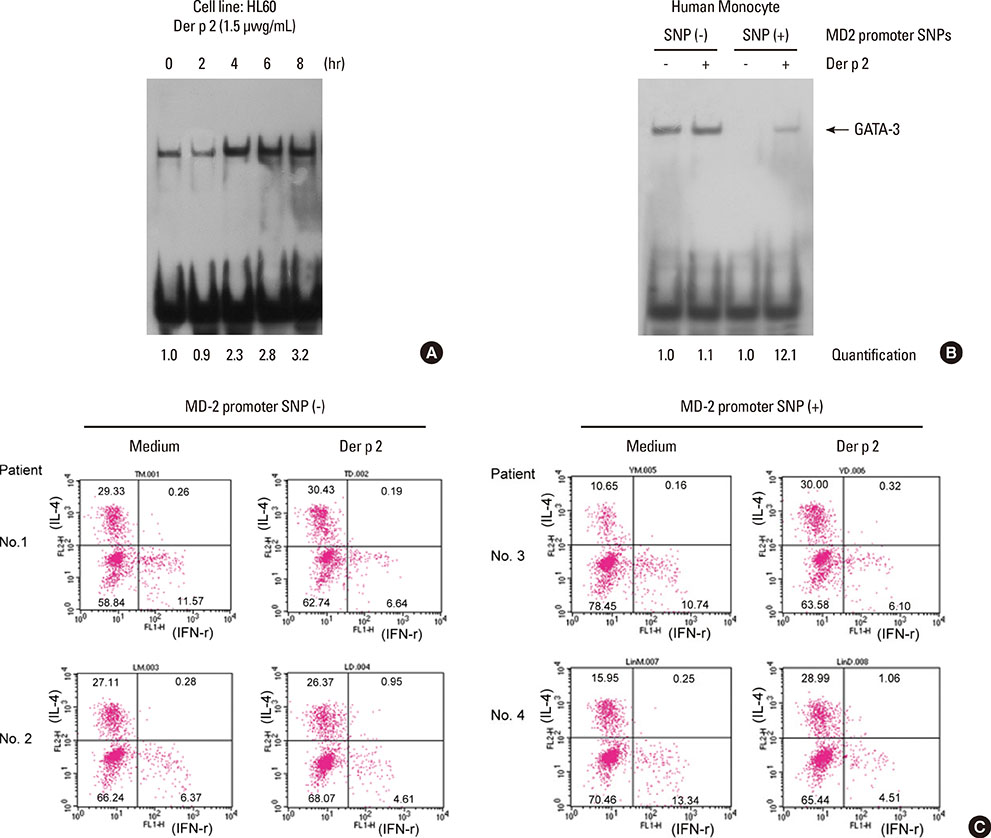
Fig. 6 MD-2 promoter SNPs (rs1809441/rs1809442) affect the binding affinity of GATA-3 and TH2-cytokine expression. The binding activity of GATA-3 and TH2-cytokine expression were analyzed by electrophoretic mobility shift assay (EMSA) and flow cytometry, respectively. The human promyelocytic leukemia cell line (HL-60) and human monocyte derived from donor' PBMCs were employed for GATA-3 binding activity. The binding activity of GATA-3 to these MD-2 promoter SNPs was analyzed by EMSA using an oligonucleotide containing the tandem GATA motif (-136 to -161). (A) The binding activity of the transcription factor GATA-3 was induced by Der p 2 (1.5 µg/mL) at different time points (0-8 hours) with the human promyelocytic leukemia cell line HL-60. (B) Human monocytes derived from subjects with (+) or without (-) MD-2 promoter SNPs were collected to analyze the binding affinity of GATA-3. SNP (+), MD-2 promoter SNPs (rs1809441-T/rs1809442-G); SNP (-), MD-2 promoter SNPs (rs1809441-G /rs1809442-C). Quantification: hybridization signals were quantified by the image analysis program. (C) Cytokines in activated TH cells derived from donor's leukocytes underwent immunofluorescence staining and flow cytometry. Three-color staining methods were used to analyze the expressions of IL-4 and interferon (IFN)-γ in CD4+ cells. Mean fluorescence was measured using a flow cytometer and is presented in percentages. A total of 5,000 cells were analyzed in each sample.
Reference
-
1. Tovey ER, Chapman MD, Platts-Mills TA. Mite faeces are a major source of house dust allergens. Nature. 1981; 289:592–593.2. Boulet LP, Turcotte H, Laprise C, Lavertu C, Bédard PM, Lavoie A, et al. Comparative degree and type of sensitization to common indoor and outdoor allergens in subjects with allergic rhinitis and/or asthma. Clin Exp Allergy. 1997; 27:52–59.3. Thomas WR, Smith WA, Hales BJ, Mills KL, O'Brien RM. Characterization and immunobiology of house dust mite allergens. Int Arch Allergy Immunol. 2002; 129:1–18.4. Tsai JJ, Shen HD, Chua KY. Purification of group 2 Dermatophagoides pteronyssinus allergen and prevalence of its specific IgE in asthmatics. Int Arch Allergy Immunol. 2000; 121:205–210.5. Shin JW, Sue JH, Song TW, Kim KW, Kim ES, Sohn MH, et al. Atopy and house dust mite sensitization as risk factors for asthma in children. Yonsei Med J. 2005; 46:629–634.6. Park SH, Kim ND, Jung JK, Lee CK, Han SB, Kim Y. Myeloid differentiation 2 as a therapeutic target of inflammatory disorders. Pharmacol Ther. 2012; 133:291–298.7. Ohto U, Fukase K, Miyake K, Satow Y. Crystal structures of human MD-2 and its complex with antiendotoxic lipid IVa. Science. 2007; 316:1632–1634.8. Trompette A, Divanovic S, Visintin A, Blanchard C, Hegde RS, Madan R, et al. Allergenicity resulting from functional mimicry of a Toll-like receptor complex protein. Nature. 2009; 457:585–588.9. Vasl J, Prohinar P, Gioannini TL, Weiss JP, Jerala R. Functional activity of MD-2 polymorphic variant is significantly different in soluble and TLR4-bound forms: decreased endotoxin binding by G56R MD-2 and its rescue by TLR4 ectodomain. J Immunol. 2008; 180:6107–6115.10. Tsai JJ, Liu SH, Yin SC, Yang CN, Hsu HS, Chen WB, et al. Mite allergen Der-p2 triggers human B lymphocyte activation and Toll-like receptor-4 induction. PLoS ONE. 2011; 6:e23249.11. Hasegawa M, Nishiyama C, Nishiyama M, Akizawa Y, Mitsuishi K, Ito T, et al. A novel -66T/C polymorphism in Fc epsilon RI alpha-chain promoter affecting the transcription activity: possible relationship to allergic diseases. J Immunol. 2003; 171:1927–1933.12. Liao EC, Chang CY, Wu CC, Wang GJ, Tsai JJ. Association of single nucleotide polymorphisms in the MD-2 gene promoter region with Der p 2 allergy. Allergy Asthma Immunol Res. 2015; 7:249–255.13. Gu W, Shan YA, Zhou J, Jiang DP, Zhang L, Du DY, et al. Functional significance of gene polymorphisms in the promoter of myeloid differentiation-2. Ann Surg. 2007; 246:151–158.14. Bousquet J, Van Cauwenberge P, Khaltaev N. Aria Workshop Group. World Health Organization. Allergic rhinitis and its impact on asthma. J Allergy Clin Immunol. 2001; 108:S147–S334.15. Bousquet J, Khaltaev N, Cruz AA, Denburg J, Fokkens WJ, Togias A, et al. Allergic Rhinitis and its Impact on Asthma (ARIA) 2008 update (in collaboration with the World Health Organization, GA(2)LEN and AllerGen). Allergy. 2008; 63:Suppl 86. 8–160.16. Noguchi T, Matsuda H, Akiyama Y. PDB-REPRDB: a database of representative protein chains from the Protein Data Bank (PDB). Nucleic Acids Res. 2001; 29:219–220.17. Mendy A, Gasana J, Vieira ER, Forno E, Patel J, Kadam P, et al. Endotoxin exposure and childhood wheeze and asthma: a meta-analysis of observational studies. J Asthma. 2011; 48:685–693.18. Chiou YL, Lin CY. Der p2 activates airway smooth muscle cells in a TLR2/MyD88-dependent manner to induce an inflammatory response. J Cell Physiol. 2009; 220(2):311–318.19. Osterlund C, Grönlund H, Polovic N, Sundström S, Gafvelin G, Bucht A. The non-proteolytic house dust mite allergen Der p 2 induce NF-κB and MAPK dependent activation of bronchial epithelial cells. Clin Exp Allergy. 2009; 39:1199–1208.20. Zhu J, Yamane H, Cote-Sierra J, Guo L, Paul WE. GATA-3 promotes Th2 responses through three different mechanisms: induction of Th2 cytokine production, selective growth of Th2 cells and inhibition of Th1 cell-specific factors. Cell Res. 2006; 16:3–10.21. Paul WE, Zhu J. How are T(H)2-type immune responses initiated and amplified? Nat Rev Immunol. 2010; 10:225–235.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Association of Single Nucleotide Polymorphisms in the MD-2 Gene Promoter Region With Der p 2 Allergy
- Der p 1, Der p 2 and Der p 10 IgE Reactivities in Allergic Rhinitis Patients in Korea
- Immunoglobulin E to allergen components of house dust mite in Korean children with allergic disease
- Strain-Specific Differences in House Dust Mite (Dermatophagoides farinae)-Induced Mouse Models of Allergic Rhinitis
- IL-13 Gene Polymorphisms are Associated With Rhinosinusitis and Eosinophilic Inflammation in Aspirin Intolerant Asthma
