Immunohistochemical study on the expression of calcium binding proteins (calbindin-D28k, calretinin, and parvalbumin) in the cerebral cortex and in the hippocampal region of nNOS knock-out(-/-) mice
- Affiliations
-
- 1Department of Anatomy, College of Medicine, Seoul National University, Seoul, Korea. kmjoo@snu.ac.kr
- 2Department of Neurosurgery, School of Medicine, Sungkyunkwan University, Seoul, Korea.
- KMID: 1447420
- DOI: http://doi.org/10.5115/acb.2011.44.2.106
Abstract
- Nitric oxide (NO) modulates the activities of various channels and receptors to participate in the regulation of neuronal intracellular Ca2+ levels. Ca2+ binding protein (CaBP) expression may also be altered by NO. Accordingly, we examined expression changes in calbindin-D28k, calretinin, and parvalbumin in the cerebral cortex and hippocampal region of neuronal NO synthase knockout(-/-) (nNOS-/-) mice using immunohistochemistry. For the first time, we demonstrate that the expression of CaBPs is specifically altered in the cerebral cortex and hippocampal region of nNOS-/- mice and that their expression changed according to neuronal type. As changes in CaBP expression can influence temporal and spatial intracellular Ca2+ levels, it appears that NO may be involved in various functions, such as modulating neuronal Ca2+ homeostasis, regulating synaptic transmission, and neuroprotection, by influencing the expression of CaBPs. Therefore, these results suggest another mechanism by which NO participates in the regulation of neuronal Ca2+ homeostasis. However, the exact mechanisms of this regulation and its functional significance require further investigation.
Keyword
MeSH Terms
-
Animals
Calcium
Calcium-Binding Protein, Vitamin D-Dependent
Calcium-Binding Proteins
Carrier Proteins
Cerebral Cortex
Homeostasis
Immunohistochemistry
Mice
Neurons
Nitric Oxide
Nitric Oxide Synthase
Synaptic Transmission
Calcium
Calcium-Binding Protein, Vitamin D-Dependent
Calcium-Binding Proteins
Carrier Proteins
Nitric Oxide
Nitric Oxide Synthase
Figure
-

Fig. 1 Changes in calbindin D28k (CB) immunoreactivity in the cerebral cortex of neuronal NO synthase knockout(-/-) (nNOS-/-) mice (D-F) compared with control mice (A-C). (B, E) are high power views of layers II and III and (C, F) are high power views of layer V of the cerebral cortex. The distribution pattern and morphology of CB-immunoreactive neurons were similar in the control and nNOS-/- mice (A, D). CB immunoreactivity in nNOS-/- mice was much lower than that of control mice (D-F) and the high power views show that lengths of the CB-immunoreactive neurites in the nNOS-/- mice were shorter and less branched than those of the control mice (black arrowheads in E and F). Scale bars=200 µm (A, D), 50 µm (B, C, E, F).
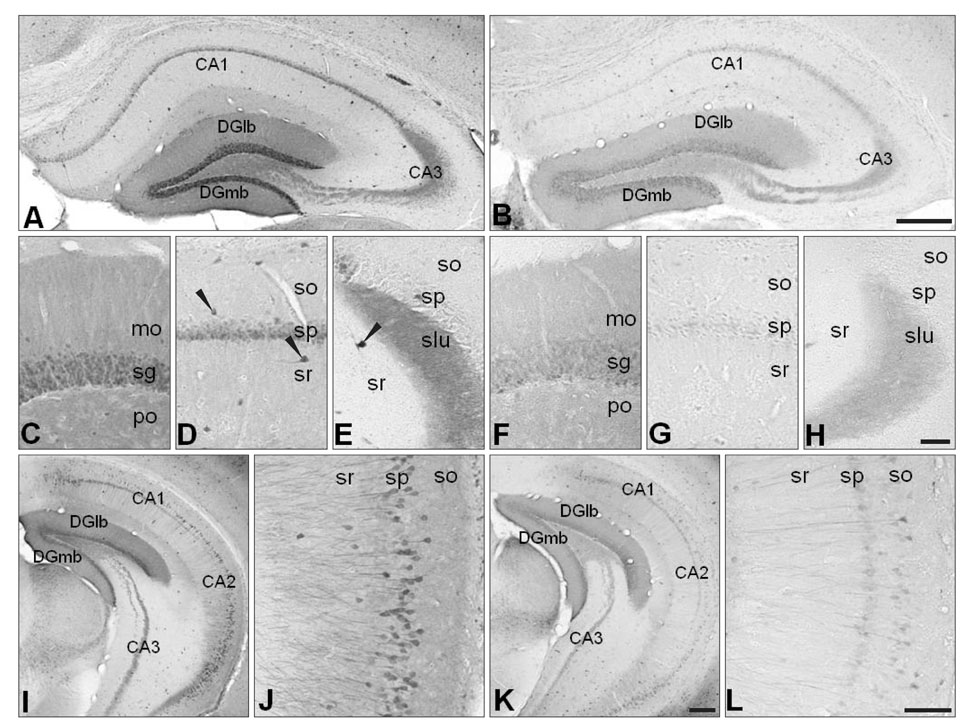
Fig. 2 Decreased calbindin D28k (CB) expression in the hippocampal region of neuronal NO synthase knockout(-/-) (nNOS-/-) mice (A, C-E, I, J for the control mice and B, F-H, K, L for the nNOS-/- mice). (C and F, D and G, and E and H) are high power views of the dentate gyrus, CA1 of Ammon's horn, and CA3 of Ammon's horn, respectively. The overall distribution patterns of CB-immunoreactivity and the morphology of CB-immunoreactive neurons were similar in the control and nNOS-/- mice (A-L). CB immunoreactivity was much reduced in the neuronal cell bodies and the neuropil of nNOS-/- mice (B, F-H, K, L). CA1, field CA1 Ammon's horn; CA2, field CA2 Ammon's horn; CA3, field CA3 Ammon's horn; DGlb, dentate gyrus lateral blade; DGmb, dentate gyrus medial blade; mo, molecular layer; sg, granule cell layer; po, polymorph layer; so, stratum oriens; sp, pyramidal layer; sr, stratum radiatum; slu, stratum lucidum. Scale bars=200 µm (A, B), 50 µm (C-H), 100 µm (I, K), 80 µm (J, L).
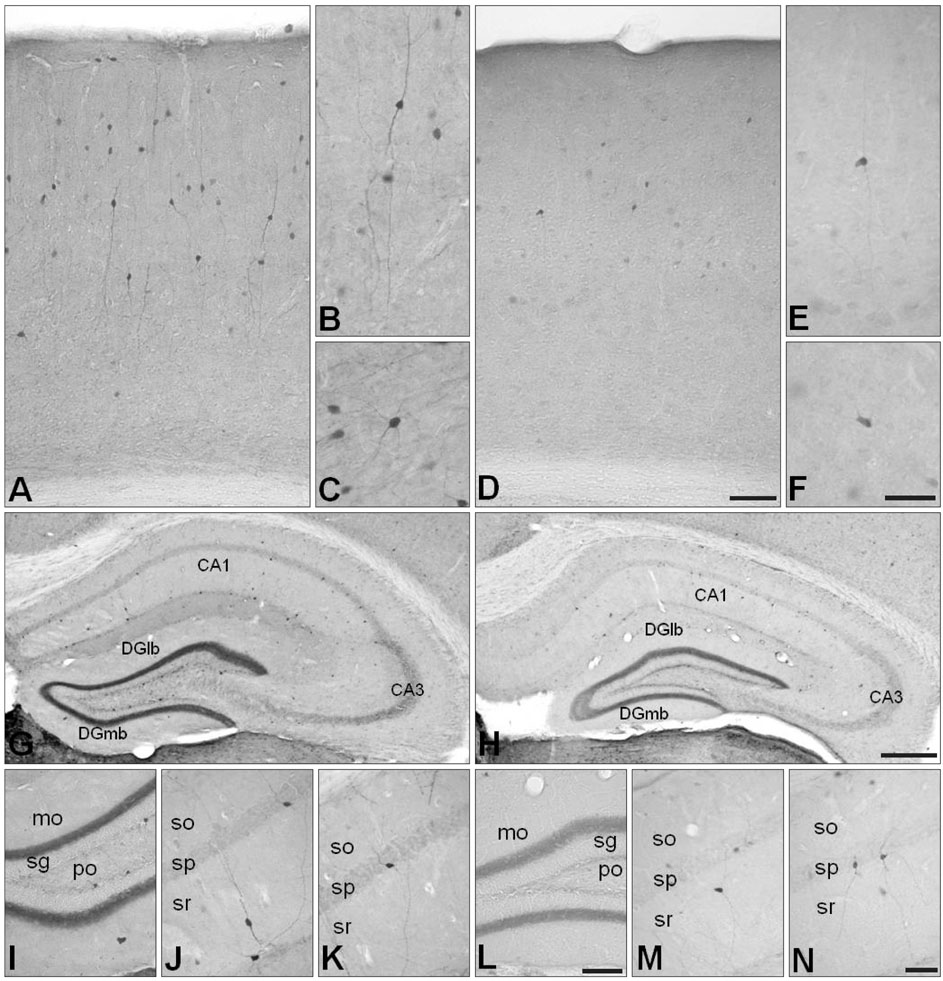
Fig. 3 Changes in calretinin (CR) immunoreactivity in the cerebral cortex (D-F) and in the hippocampal region (H, L-N) of neuronal NO synthase knockout(-/-) (nNOS-/-) mice, compared with those of control mice (A-C for the cerebral cortex and G, I-K for the hippocampal region). (I and L, J and K, and M and N) are high power views of the dentate gyrus and the CA1 of Ammon's horn, respectively. In the cerebral cortex and hippocampal region, the overall distribution patterns of CR-immunoreactivity and the morphology of CR-immunoreactive neurons were similar in the control and nNOS-/- mice (A-N). In the cerebral cortex, nNOS-/- mice had fewer CR-immunoreactive neurons and each CR-immunoreactive neuron showed less CR-immunoreactivity than that in control mice (D-F). High power views showed that the lengths of the CR-immunoreactive neurites in the nNOS-/- mice were shorter than those of the control mice (D-F). Calbindin D28k (CB) immunoreactivity in the neuropil of the dentate gyrus was much reduced in the hippocampal region of nNOS-/- mice (H, L) and the number of CR-immunoreactive neurons was less (H) in nNOS-/- mice, although the morphology of these neurons was preserved (L-N). CA1, field CA1 Ammon's horn; CA3, field CA3 Ammon's horn; DGlb, dentate gyrus lateral blade; DGmb, dentate gyrus medial blade; mo, molecular layer; sg, granule cell layer; po, polymorph layer; so, stratum oriens; sp, pyramidal layer; sr, stratum radiatum. Scale bars=150 µm (A, D), 80 µm (B, C, E, F), 200 µm (G, H), 65 µm (I, L), 50 µm (J, K, M, N).

Fig. 4 Parvalbumin (PV) immunoreactivity in the cerebral cortex (A-D) and hippocampal region (E-L) of control mice (A, B, E, G-I) and neuronal NO synthase knockout(-/-) (nNOS-/-) mice (C, D, F, J-L). (B and C, G and J, H and K, and I and L) are high power views of the cerebral cortex, the polymorph layer of the dentate gyrus, the granule cell layer of the dentate gyrus, and the pyramidal layer of Ammon's horn, respectively. The numbers of PV-immunoreactive neurons, their distribution patterns, and morphology were unchanged in the cerebral cortex and hippocampal region of nNOS-/- mice (F, J-L). CA1, field CA1 Ammon's horn; CA3, field CA3 Ammon's horn; DGlb, dentate gyrus lateral blade; DGmb, dentate gyrus medial blade; sg, granule cell layer; po, polymorph layer; mo, molecular layer;so, stratum oriens; sp, pyramidal layer; sr, stratum radiatum. Scale bars=150 µm (A, D), 60 µm (B, C), 200 µm (E, F), 40 µm (G-L).
Reference
-
1. Moncada S, Higgs A. The L-arginine-nitric oxide pathway. N Engl J Med. 1993. 329:2002–2012.2. Liu PK, Robertson CS, Valadka A. The association between neuronal nitric oxide synthase and neuronal sensitivity in the brain aft er brain injury. Ann N Y Acad Sci. 2002. 962:226–241.3. Clementi E, Riccio M, Sciorati C, Nisticò G, Meldolesi J. The type 2 ryanodine receptor of neurosecretory PC12 cells is activated by cyclic ADP-ribose. Role of the nitric oxide/cGMP pathway. J Biol Chem. 1996. 271:17739–17745.4. Short DM, Heron ID, Birse-Archbold JL, Kerr LE, Sharkey J, McCulloch J. Apoptosis induced by staurosporine alters chaperone and endoplasmic reticulum proteins: identification by quantitative proteomics. Proteomics. 2007. 7:3085–3096.5. Mattson MP. Apoptosis in neurodegenerative disorders. Nat Rev Mol Cell Biol. 2000. 1:120–129.6. Rogers JH. Calretinin: a gene for a novel calcium-binding protein expressed principally in neurons. J Cell Biol. 1987. 105:1343–1353.7. Polans A, Baehr W, Palczewski K. Turned on by Ca2+! The physiology and pathology of Ca(2+)-binding proteins in the retina. Trends Neurosci. 1996. 19:547–554.8. Schäfer BW, Heizmann CW. The S100 family of EF-hand calcium-binding proteins: functions and pathology. Trends Biochem Sci. 1996. 21:134–140.9. Geula C, Schatz CR, Mesulam MM. Differential localization of NADPH-diaphorase and calbindin-D28k within the cholinergic neurons of the basal forebrain, striatum and brainstem in the rat, monkey, baboon and human. Neuroscience. 1993. 54:461–476.10. Bertini G, Peng ZC, Bentivoglio M. The chemical heterogeneity of cortical interneurons: nitric oxide synthase vs. calbindin and parvalbumin immunoreactivity in the rat. Brain Res Bull. 1996. 39:261–266.11. Arévalo R, Sánchez F, Alonso JR, Rubio M, Aijón J, Vázquez R. Infrequent cellular coexistence of NADPH-diaphorase and calretinin in the neurosecretory nuclei and adjacent areas of the rat hypothalamus. J Chem Neuroanat. 1993. 6:335–341.12. Nelson RJ, Demas GE, Huang PL, Fishman MC, Dawson VL, Dawson TM, Snyder SH. Behavioural abnormalities in male mice lacking neuronal nitric oxide synthase. Nature. 1995. 378:383–386.13. Airaksinen MS, Eilers J, Garaschuk O, Thoenen H, Konnerth A, Meyer M. Ataxia and altered dendritic calcium signaling in mice carrying a targeted null mutation of the calbindin D28k gene. Proc Natl Acad Sci U S A. 1997. 94:1488–1493.14. Cheron G, Schurmans S, Lohof A, d'Alcantara P, Meyer M, Draye JP, Parmentier M, Schiffmann SN. Electrophysiological behavior of Purkinje cells and motor coordination in calretinin knock-out mice. Prog Brain Res. 2000. 124:299–308.15. Huang PL, Dawson TM, Bredt DS, Snyder SH, Fishman MC. Targeted disruption of the neuronal nitric oxide synthase gene. Cell. 1993. 75:1273–1286.16. Baimbridge KG, Celio MR, Rogers JH. Calcium-binding proteins in the nervous system. Trends Neurosci. 1992. 15:303–308.17. Freund TF, Buzsáki G. Interneurons of the hippocampus. Hippocampus. 1996. 6:347–470.18. Kawaguchi Y, Kubota Y. GABAergic cell subtypes and their synaptic connections in rat frontal cortex. Cereb Cortex. 1997. 7:476–486.19. Lee JC, Cho YJ, Kim J, Kim N, Kang BG, Cha CI, Joo KM. Region-specific changes in the immunoreactivity of vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide receptors (VPAC2, and PAC1 receptor) in the aged rat brains. Brain Res. 2010. 1351:32–40.20. DeFelipe J. Types of neurons, synaptic connections and chemical characteristics of cells immunoreactive for calbindin-D28K, parvalbumin and calretinin in the neocortex. J Chem Neuroanat. 1997. 14:1–19.21. Kawaguchi Y, Kondo S. Parvalbumin, somatostatin and chole cy stokinin as chemical markers for specific GABAergic interneuron types in the rat frontal cortex. J Neurocytol. 2002. 31:277–287.22. Jinno S, Kosaka T. Patterns of expression of calcium binding proteins and neuronal nitric oxide synthase in different populations of hippocampal GABAergic neurons in mice. J Comp Neurol. 2002. 449:1–25.23. Jouvenceau A, Potier B, Battini R, Ferrari S, Dutar P, Billard JM. Glutamatergic synaptic responses and long-term potentiation are impaired in the CA1 hippocampal area of calbindin D(28k)-deficient mice. Synapse. 1999. 33:172–180.24. Schurmans S, Schiffmann SN, Gurden H, Lemaire M, Lipp HP, Schwam V, Pochet R, Imperato A, Böhme GA, Parmentier M. Impaired long-term potentiation induction in dentate gyrus of calretinin-deficient mice. Proc Natl Acad Sci U S A. 1997. 94:10415–10420.25. Parra P, Gulyás AI, Miles R. How many subtypes of inhibitory cells in the hippocampus? Neuron. 1998. 20:983–993.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Immunohistochemical study on the expression of calcium binding proteins (calbindin-D28k, calretinin, and parvalbumin) in the cerebellum of the nNOS knock-out(-/-) mice
- Seizure -Related Change of NADPH -diaphorase and Calcium Binding Protein Positive Neurons in the Brain of Rats
- The distribution of calbindin-D28k, parvalbumin, and calretinin immunoreactivity in the inferior colliculus of circling mouse
- Immunocytochemical Studies of Calbindin D-28k and Parvalbumin in the Sensory and motor Cortex of the Cat
- Changes in transcript and protein levels of calbindin D28k, calretinin and parvalbumin, and numbers of neuronal populations expressing these proteins in an ischemia model of rat retina
