Enhancement of Plasmacytoma Cell Growth by Ascorbic Acid is Mediated Via Glucose 6-phosphate Dehydrogenase
- Affiliations
-
- 1Korea Institute of Toxicology, Daejeon, Korea. wskoh@kitox.re.kr
- 2Cancer Center, Samsung Medical Center, Sungkyunkwan University School of Medicine, Seoul, Korea.
- 3The Center for Improvement of Human Functioning International, Wichita, KS 67219, USA.
Abstract
-
PURPOSE: We investigated the mechanism by which some types of cancer cells grow faster in the presence of ascorbic acid supplementation.
MATERIALS AND METHODS
Adj.PC-5, a mouse plasmacytoma cell, is known to show ascorbic acid-dependent growth and was chosen as a test system. The growth of cancer cells was measured by the colony number on soft agar or the cellular proliferation in suspension culture. The ascorbate level was measured by a high performance liquid chromatography system with an electrochemical detector. Glucose 6-phosphate dehydrogenase was analyzed both on the specific enzyme activity level and on the transcription level by performing Northern blot analysis.
RESULTS
Ascorbyl 2-phosphate among the ascorbate derivatives was the most efficient in stimulating cell growth. The intracellular and extracellular ascorbate concentrations following treatment with either ascorbate or ascorbyl 2-phosphate suggest that the superiority of ascorbyl 2-phosphate for stimulating cell growth may be due to its slow conversion to ascorbate in the culture medium. The steady transformation to ascorbate ensures sustained levels of ascorbate in the culture medium and thereby maximizes the growth stimulatory effect of ascorbate. Ascorbyl 2-phosphate markedly enhanced, in a concentration-and time-dependent manner, mRNA synthesis as well as the enzymatic activity of glucose 6- phosphate dehydrogenase, which is known to be a rate- limiting enzyme in cell growth. On the other hand, simultaneous addition of dehydroisoandrosterone, a well- known inhibitor of glucose 6-phosphate dehydrogenase, to the culture medium abrogated the growth stimulation by ascorbyl 2-phosphate, and it also reduced the glucose 6-phosphate dehydrogenase activity proportionately.
CONCLUSIONS
The results from this study suggest that enhanced glucose 6-phosphate dehydrogenase activity may at least in part explain the stimulation of cell growth by ascorbate or ascorbyl 2-phosphate.
MeSH Terms
Figure
-
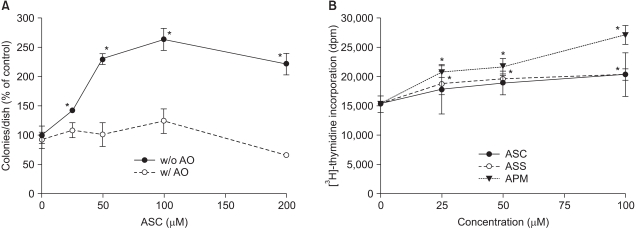
Fig. 1 Ascorbate-induced stimulation of Adj.PC-5 cell growth. (A) Colony growth in the Adj.PC-5 cell cultures in the presence (open circles) or absence (closed circles) of ascorbate oxidase (AO). Spleen cells (3×105 nucleated cells/dish) from the plasmacytoma (Adj.PC-5)-infiltrated spleens of BALB/c mice were cultured on soft-agar (0.3%) plates. The cultures were daily fed with 0.5 ml of complete RPMI 1640 medium with ascorbate in the presence or absence of ascorbate oxidase (0.5 unit/ml). The colony counts were normalized to the untreated control (52 colonies/dish). (B) Effects of ascorbate derivatives on the proliferation of Adj.PC-5 cells. Ascorbate derivatives were directly added to cell suspensions (3×105 nucleated cells/ml), which were then transferred in 200µl aliquots and cultured on 96-well microtiter plates at 37℃ and 5% CO2 for 48 h. Cell growth was determined by the incorporation of [3H]-thymidine for the last 6 h of the culture period. Each point shows the mean±SD of quadruplicate determinations. *significantly different from control (p<0.05). ASC, ascorbic acid (closed circles); ASS, sodium ascorbate (open circles); APM, magnesium ascorbyl 2-phosphate (closed triangles).
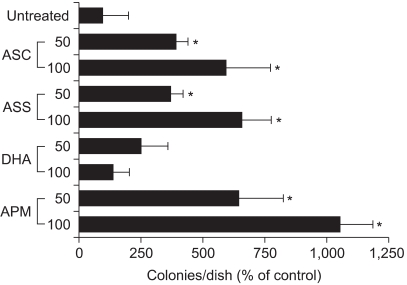
Fig. 2 Effects of the various forms of ascorbate on colony growth in Adj.PC-5 cell cultures. Adj.PC-5-infiltrated spleen cells were adjusted to 3×105 nucleated cells/dish and then cultured as described in Fig. 1. The cultures were daily fed with 0.5 ml of complete RPMI 1640 medium containing either 50 or 100µM of the indicated ascorbate derivative. The colony counts were normalized to the untreated control. Each bar represents the mean±SD of triplicate determinations. *significantly different from control (p<0.05). DHA, dehydroascorbic acid.
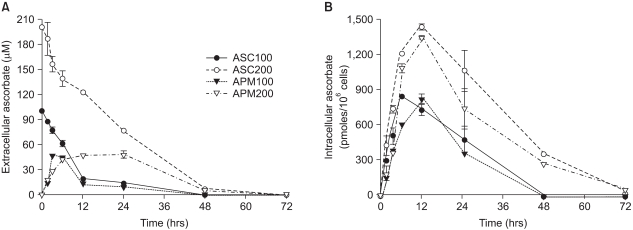
Fig. 3 Differential kinetics of ascorbate and APM in the cells and the culture medium. Prior to the experiments, Adj.PC-5 cells were maintained in medium that contained no ascorbate for at least 10 days to confirm they had no endogenous ascorbate. Experimental cultures were treated with either 100µM (closed circles) or 200µM (open circles) of ascorbate or either 100µM (closed triangles) or 200µM (open triangles) of APM for the indicated times and then the cells and supernatants separately collected. (A) Extracellular ascorbate concentration. The culture supernatant was mixed with an equal volume of metaphosphoric acid (10%) to extract the ascorbic acid. (B) Intracellular ascorbate concentration. The cells were washed with phosphate-buffered saline and extracted in 5% metaphosphoric acid solution for 10 min. The extracts from both the cells and culture supernatants were filtered (pore size: 0.22µm) and then applied to the HPLC analysis as described in Materials and Methods. Each point represents the mean±SD of triplicate determinations.
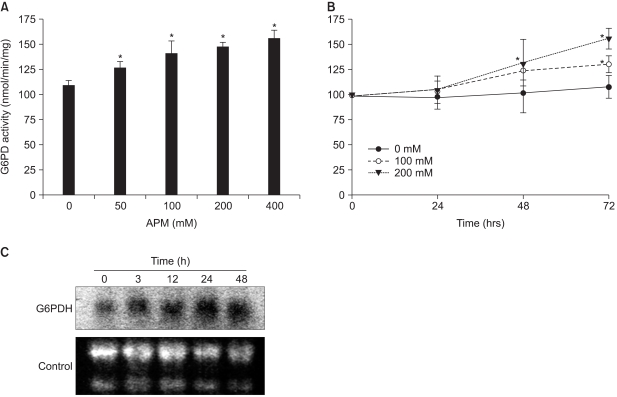
Fig. 4 Augmented G6PD activity and mRNA in the Adj.PC-5 cells cultured in the presence of APM. (A) The G6PD activity increase induced by APM was concentration dependent. Adj.PC-5 cells (approximately 5×106 cells) were harvested after 72 h incubation with APM and then homogenized. The supernatants after centrifugation at 14,000 rpm for 10 min were used for G6PD assays. (B) Time course of the increased G6PD activity. G6PD activities were measured from the Adj.PC-5 cells incubated with 0 (closed circles), 100 (open circles), or 200 (closed triangles)µM of APM for the indicated times. G6PD activity was measured by spectrophotometry at 340 nm by the reduction of NADP+ in the presence of glucose-6-phosphate. Each point represents the mean activity±SD of triplicate determinations. (C) Northern blot analysis of the G6PD mRNA. The total RNA was isolated from the cells treated with APM (200µM) for 0, 3, 12, 24 and 48 h, respectively. The RNA (30µg) was electrophoresed, transferred to nylon membranes and hybridized with a 32P-labeled probe of mouse G6PD cDNA. The control, 28S and 18S RNA in the gel.
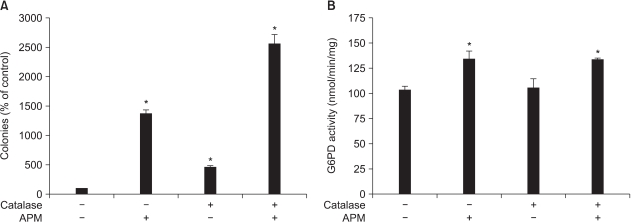
Fig. 5 Effect of adding catalase to the Adj.PC-5 cell cultures. (A) Colony growth stimulated by APM. Colony forming assay was performed as described in Materials and Methods. Catalase and APM were prepared in the feeding medium and this was added daily to culture. (B) G6PD activity enhanced by APM. The cells were cultured with the indicated treatments for 72 h and then the cells were collected. G6PD activity was determined as described in Materials and Methods. Each bar represents the mean±SD of triplicate determinations. *significantly different from control (p<0.05). Catalase (10 units/ml); APM (200µM).
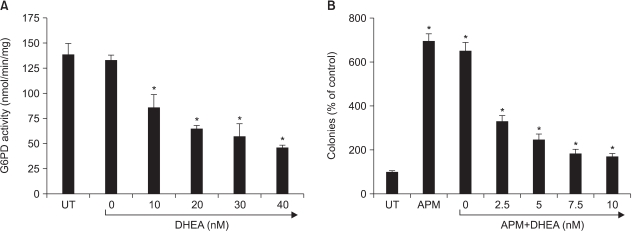
Fig. 6 Effect of DHEA. (A) Effect on G6PD activity in Adj.PC5 cell lysates. The cell lysate for G6PD assay was prepared as described in Materials and Methods. DHEA was tested to determine its inhibitory effect on G6PD activity. Each point represents the mean±SD of triplicate determinations. The effect of DHEA, dehydroepiandrosterone, on colony growth enhanced by the addition of APM. The colony forming assay was performed as described in Materials and Methods. DHEA was dissolved in dimethylsulfoxide (final dimethylsulfoxide concentration in medium: 0.1%) and this was added daily to the Adj.PC-5 cell cultures that were also treated with APM. Each bar represents the mean±SD of quadruplicate cultures. *significantly different from control (p<0.05).
Reference
-
1. Elmore AR. Final report of the safety assessment of L-Ascorbic Acid, Calcium Ascorbate, Magnesium Ascorbate, Magnesium Ascorbyl Phosphate, Sodium Ascorbate, and Sodium Ascorbyl Phosphate as used in cosmetics. Int J Toxicol. 2005; 24(Suppl 2):51–111. PMID: 16154915.2. Casciari JJ, Riordan NH, Schmidt TL, Meng XL, Jackson JA, Riordan HD. Cytotoxicity of ascorbate, lipoic acid, and other antioxidants in hollow fibre in vitro tumours. Br J Cancer. 2001; 84:1544–1550. PMID: 11384106.
Article3. Koh WS, Lee SJ, Lee H, Park C, Park MH, Kim WS, et al. Differential effects and transport kinetics of ascorbate derivatives in leukemic cell lines. Anticancer Res. 1998; 18:2487–2493. PMID: 9703897.4. Park CH, Amare M, Hoogstraten B. Analysis of the growth enhancing effect of L-ascorbic acid on human leukemic cells in culture. Exp Hematol. 1980; 8:853–859. PMID: 16398016.5. Park CH, Bergsagel DE, McCulloch EA. Ascorbic acid: a culture requirement for colony formation by mouse plasmacytoma cells. Science. 1971; 174:720–722. PMID: 5123422.
Article6. Venezian R, Shenker BJ, Datar S, Leboy PS. Modulation of chondrocyte proliferation by ascorbic acid and BMP-2. J Cell Physiol. 1998; 174:331–341. PMID: 9462695.
Article7. Nowak G, Schnellmann RG. L-ascorbic acid regulates growth and metabolism of renal cells: improvements in cell culture. Am J Physiol. 1996; 271:C2072–C2080. PMID: 8997210.
Article8. Deutsch JC. Ascorbic acid oxidation by hydrogen peroxide. Anal Biochem. 1998; 255:1–7. PMID: 9448835.
Article9. Mendiratta S, Qu Z, May JM. Erythrocyte defenses against hydrogen peroxide: the role of ascorbic acid. Biochim Biophys Acta. 1998; 1380:389–395. PMID: 9555101.
Article10. Yoshimoto K, Nakamura T, Ichihara A. Reciprocal effects of epidermal growth factor on key lipogenic enzymes in primary cultures of adult rat hepatocytes. Induction of glucose-6-phosphate dehydrogenase and suppression of malic enzyme and lipogenesis. J Biol Chem. 1983; 258:12355–12360. PMID: 6355085.
Article11. Baba M, Yamamoto R, Iishi H, Tatsuta M, Wada A. Role of glucose-6-phosphate dehydrogenase on enhanced proliferation of pre-neoplastic and neoplastic cells in rat liver induced by N-nitrosomorpholine. Int J Cancer. 1989; 43:892–895. PMID: 2565888.
Article12. Deshpande N, Mitchell I, Millis R. Enzyme studies in human breast tumours. Eur J Cancer. 1977; 13:1261–1267. PMID: 145367.
Article13. Heyden G. Histochemical investigation of malignant cells. Histochemistry. 1974; 39:327–334. PMID: 4859353.
Article14. Shepherd A, Cleary MP. Metabolic alterations after dehydroepiandrosterone treatment in Zucker rats. Am J Physiol. 1984; 246:E123–E128. PMID: 6230014.
Article15. Tian WN, Braunstein LD, Pang J, Stuhlmeier KM, Xi QC, Tian X, et al. Importance of glucose-6-phosphate dehydrogenase activity for cell growth. J Biol Chem. 1998; 273:10609–10617. PMID: 9553122.
Article16. Park CH, Bergsagel DE, McCulloch EA. Mouse myeloma tumor stem cells: a primary cell culture assay. J Natl Cancer Inst. 1971; 46:411–422. PMID: 5115909.17. Kornberg A, Horecker BL. Colowick SP, Kaplan NO, editors. Glucose-6-phosphate dehydrogenase. Methods of enzymology. 1955. New York: Academic Press;p. 323.18. Dunnett CW. A multiple comparison procedure for comparing several treatments with a control. J Am Statist Assoc. 1955; 50:1096–1121.
Article19. Arakawa N, Nemoto S, Suzuki E, Otsuka M. Role of hydrogen peroxide in the inhibitory effect of ascorbate on cell growth. J Nutr Sci Vitaminol (Tokyo). 1994; 40:219–227. PMID: 7965211.
Article20. Miwa N, Yamazaki H, Nagaoka Y, Kageyama K, Onoyama Y, Matsui-Yuasa I, et al. Altered production of the active oxygen species is involved in enhanced cytotoxic action of acylated derivatives of ascorbate to tumor cells. Biochim Biophys Acta. 1988; 972:144–151. PMID: 3191161.
Article21. May JM. Ascorbate function and metabolism in the human erythrocyte. Front Biosci. 1998; 3:d1–d10. PMID: 9405334.
Article22. Kuo SM, MacLean ME, McCormick K, Wilson JX. Gender and sodium-ascorbate transporter isoforms determine ascorbate concentrations in mice. J Nutr. 2004; 134:2216–2221. PMID: 15333707.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Jaundice and Hemolytic Anemia Appearing within the First 24 Hour of Life due to Glucose-6-Phosphate Dehydrogenase Deficiency
- The Effect of Ascorbic Acid and its Derivative on Cultured Rabbit Keratocytes
- A Case of Glucose-6-phosphate dehydrogenase Riley Causing Hemolytic Anemia
- Ascorbic acid insufficiency induces the severe defect on bone formation via the down-regulation of osteocalcin production
- Effects of Topically Applied Na-Hyaluronan on Epithelial Healing and Aqueous Composition in Experimental Corneal Alkali Wounds
