Effect on Cell Cycle Progression by N-Myc Knockdown in SK-N-BE(2) Neuroblastoma Cell Line and Cytotoxicity with STI-571 Compound
- Affiliations
-
- 1Department of Microbiology, Ewha Womans University School of Medicine, Seoul, Korea. soyounwoo@ewha.ac.kr
- 2Department of Pediatrics, Ewha Womans University School of Medicine, Seoul, Korea.
Abstract
-
PURPOSE: Neuroblastoma is a common tumor in childhood, and generally exhibits heterogeneity and a malignant progression. MYCN expression and amplification profiles frequently correlate with therapeutic prognosis. Although it has been reported that MYCN silencing causes differentiation and apoptosis in human neuroblastoma cells, MYCN expression influences the cytotoxic potential of chemotherapeutic drugs via the deregulation of the cell cycle. STI-571 may constitute a promising therapeutic agent against neuroblastoma, particularly in cases in which c-Kit is expressed preferentially in MYCN-amplified neuroblastoma.
MATERIALS AND METHODS
To determine whether STI-571 exerts a synergistic effect on cytotoxicity with MYCN expression, we assessed apoptotic cell death and cell cycle distribution after 72 h of exposure to STI-571 with or with out treatment of SK-N-BE(2) neuroblastoma cells with MYCN siRNA.
RESULTS
MYCN siRNA-treated SK-N-BE(2) cells did not affect apoptosis and cells were arrested in G0/G1 phase after STI-571 treatment.
CONCLUSIONS
siRNA therapy targeted to MYCN may not be effective when administered in combination with STI-571 treatment in cases of neuroblastoma. Therefore, chemotherapeutic drugs that target S or G2-M phase may prove ineffective when applied to cells arrested in the G0/1 phase as the result of MYCN knockdown and STI-571 treatment.
Keyword
MeSH Terms
Figure
-
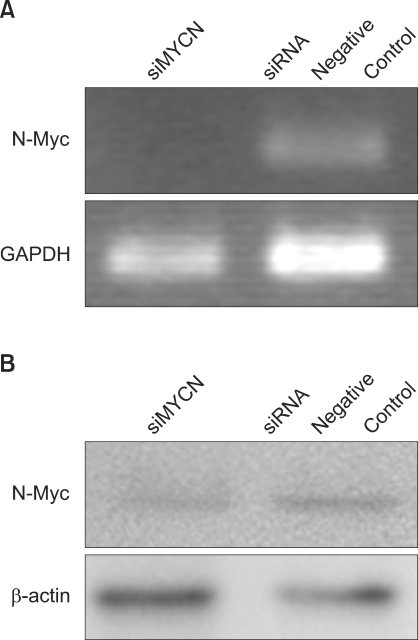
Fig. 1 Knockdown of N-myc expression using siRNA in SK-N-BE(2) cells. (A) Semi-quantitative RT-PCR analysis of MYCN mRNA expression in neuroblastoma cells after 72 h of N-Myc siRNA (siMYCN) or control siRNA transfection. GAPDH was used as a loading control. (B) Western blot analysis of N-myc protein expression in neuroblastoma cells after 72 h of transfection. β-actin was used for the equal protein loading of cell lysates. The data are expressed in arbitary pixel density units.
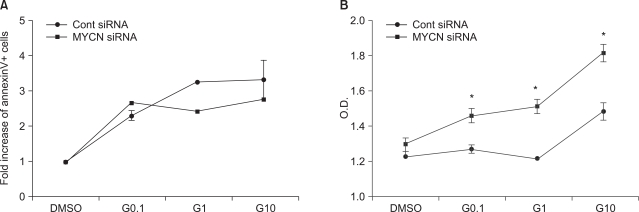
Fig. 2 Influence of STI-571 on apoptosis and proliferation. (A) AnnexinV positive and propidium iodide-negative cells were analyzed for apoptotic cells via flow cytometry with or without 0.1 uM, 1.0 uM, 10 uM of STI-571 (G0.1, G1, and G10, respectively) for 72 h. The data are expressed in terms of the fold increase in AnnexinV+ cells, expressed as the mean±SE (n=3). (B) Proliferation assay conducted in the presence of 10% FCS with or without 0.1 uM, 1.0 uM, 10 uM of STI-571 (G0.1, G1, G10, respectively) for 72 hrs. SK-N-BE(2) cells were plated at a density of 5×103/150 ul in fresh medium and treated with STI-571 as indicated. After 72 h, 20 ul of MTT (5 mg/ml in PBS) was added to each of the wells. After 4 h, the dark blue formazan crystals were dissolved, and the plates were read at a wavelength of 570 nm. The data are expressed in terms of optical density and expressed as the mean±SE (n=4). *p<0.05, NS: not statistically significant.
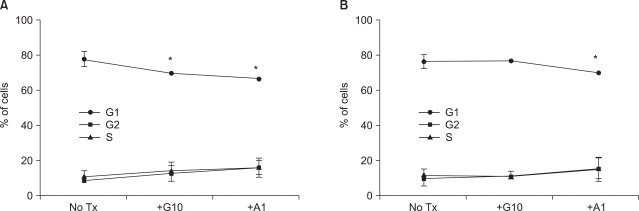
Fig. 3 MYCN knockdown by RNAi and treatment of STI-571 altered the cell cycle distribution. SK-N-BE(2) cells were transfected with control siRNA (A) or N-Myc siRNA (B) and treated with 10 uM of STI-571 or 1 uM of arsenic trioxide for 72 h. The cells were fixed with 70% ethanol followed by staining with DNA staining solution (50 ug/ml PI, 5 mM EDTA, and I mM EDTA in PBS). After collecting the data of PI positive cells via flow cytometry, cell cycles were analyzed with ModFit software for the fraction of G1 (G0/G1), G2 (G2+M), and S phases. Data are expressed as the percentage of the corresponding phase of the cells and were expressed as the mean±SE (n=4). *p<0.05.
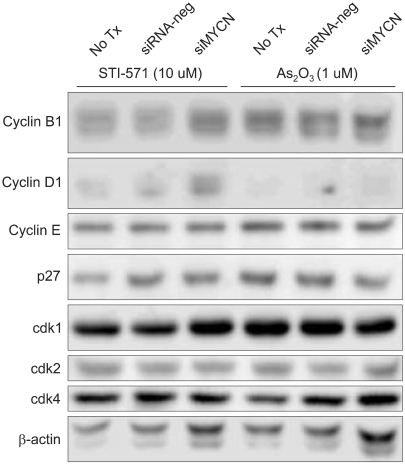
Fig. 4 Expression of cyclins, CDKs, and p27 in SK-N-BE(2) cells. (A) Cells were transfected with control siRNA (siRNA-neg) or N-Myc siRNA (siMYCN) and treated with 10 uM of STI-571 or 1 uM of arsenic trioxide for 72 h. The cells were harvested and lysed in lysis buffer containing 150 mM NaCl, 1% NP-40, 0.5% DOC, 0.1% SDS, 50 mM Tris-HCl (pH 7.5) and protease inhibitors. The protein was loaded at a concentration of 10 ug per lane on a 10% SDS-PAGE gel, transferred to a polyvinyldifluoride membrane, and immunoblotted with the indicated antibodies. β-actin blot was utilized for the equal protein loading of the cell lysates.
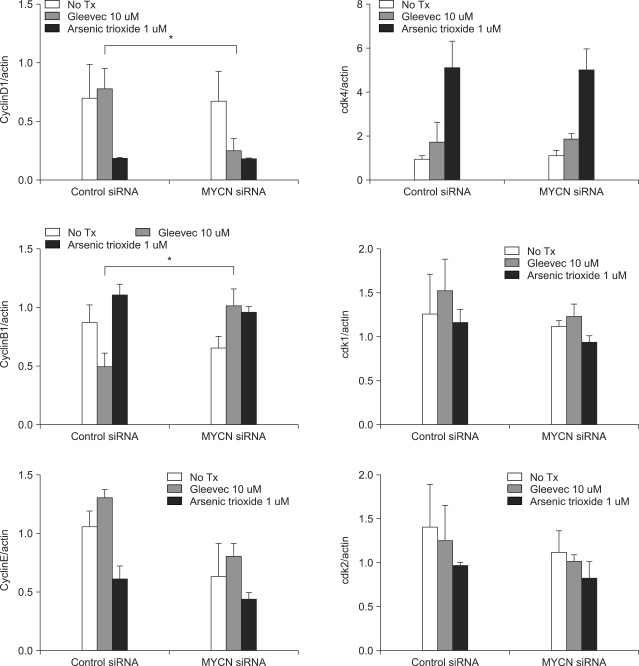
Fig. 5 Band densities were quantified by image analysis (LAS3000, Fuji, Tokyo, Japan). The relative levels of cyclins and CDKs expressed in the SK-N-BE(2) cells were corrected with the relative level of β-actin expression in the cell lysates. The mean value for the ratios of three different batches of samples is expressed. *p<0.05, NS: not statistically significant.
Reference
-
1. Kang JH, Rychahou PG, Ishola TA, Qiao J, Evers BM, Chung DH. MYCN silencing induces differentiation and apoptosis in human neuroblastoma cells. Biochem Biophys Res Commun. 2006; 351:192–197. PMID: 17055458.
Article2. Paffhausen T, Schwab M, Westermann F. Targeted MYCN expression affects cytotoxic potential of chemotherapeutic drugs in neuroblastoma cells. Cancer Lett. 2007; 250:17–24. PMID: 17141950.
Article3. Krause DS, Van Etten RA. Tyrosine kinases as targets for cancer therapy. N Engl J Med. 2005; 353:172–187. PMID: 16014887.
Article4. Mauro MJ, O'Dwyer M, Heinrich MC, Druker BJ. STI571: a paradigm of new agents for cancer therapeutics. J Clin Oncol. 2002; 20:325–334. PMID: 11773186.
Article5. Vitali R, Cesi V, Nicotra MR, McDowell HP, Donfrancesco A, Mannarino O, et al. c-Kit is preferentially expressed in MYCN-amplified neuroblastoma and its effect on cell proliferation is inhibited in vitro by STI-571. Int J Cancer. 2003; 106:147–152. PMID: 12800187.6. Meyer N, Kim SS, Penn LZ. The Oscar-worthy role of Myc in apoptosis. Semin Cancer Biol. 2006; 16:275–287. PMID: 16945552.
Article7. Schwab M, Alitalo K, Klempnauer KH, Varmus HE, Bishop JM, Gilbert F, et al. Amplified DNA with limited homology to myc cellular oncogene is shared by human neuroblastoma cell lines and a neuroblastoma tumour. Nature. 1983; 305:245–248. PMID: 6888561.
Article9. Pession A, Tonelli R. The MYCN oncogene as a specific and selective drug target for peripheral and central nervous system tumors. Curr Cancer Drug Targets. 2005; 5:273–283. PMID: 15975048.
Article10. Buchdunger E, Cioffi CL, Law N, Stover D, Ohno-Jones S, Druker BJ, et al. Abl protein-tyrosine kinase inhibitor STI571 inhibits in vitro signal transduction mediated by c-kit and platelet-derived growth factor receptors. J Pharmacol Exp Ther. 2000; 295:139–145. PMID: 10991971.11. Gilbert F, Feder M, Balaban G, Brangman D, Lurie DK, Podolsky R, et al. Human neuroblastomas and abnormalities of chromosomes 1 and 17. Cancer Res. 1984; 44:5444–5449. PMID: 6488196.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Etoposide Reduces Peroxynitrite-Induced Cytotoxicity via Direct Scavenging Effect
- The antitumor effect of various cytokines on human neuroblastoma cell lines SK-N-SH, IMR-32, and SK-N-MC
- Antiproliferative and Cytotoxic Effects of Resveratrol in Mitochondria-Mediated Apoptosis in Rat B103 Neuroblastoma Cells
- Effect of BMI1 Knockdown on Cell Proliferation, Apoptosis, Invasiveness, and Migration of U251 Glioma Cells
- Apoptosis and Cell Cycle Arrest in Two Human Breast Cancer Cell Lines by Dieckol Isolated from Ecklonia cava
