IL-8/CXCL8 Upregulates 12-Lipoxygenase Expression in Vascular Smooth Muscle Cells from Spontaneously Hypertensive Rats
- Affiliations
-
- 1Department of Microbiology, College of Medicine, Yeungnam University, Daegu, Korea. heesun@med.yu.ac.kr
- 2Department of Parmacology and Aging-associated Vascular Disease Research Center, College of Medicine, Yeungnam University, Daegu, Korea.
- KMID: 2150649
- DOI: http://doi.org/10.4110/in.2009.9.3.106
Abstract
- BACKGROUND: We previously demonstrated remarkable differences in the expression of IL-8/CXCL8 in aortic tissues and vascular smooth muscle cells (VSMC) from spontaneously hypertensive rats (SHR) compared to VSMC from normotensive Wistar-Kyoto rats (WKY). In the present study, we investigated the direct effect of IL-8/CXCL8 on expression of 12-lipoxygenase (LO), a hypertensive modulator, in SHR VSMC. METHODS: Cultured aortic VSMC from SHR and WKY were used. Expression of 12-LO mRNA was determined by real-time polymerase chain reaction. Phosphorlyation of ERK1/2 and production of 12-LO and angiotensin II subtype 1 (AT1) receptor were assessed by Western blots. IL-8/CXCL8-stimulated DNA synthesis was determined by measuring incorporation of [3H]-thymidine. And effect of IL-8/CXCL8 on vascular tone was determined by phenylephrine-induced contraction of thoracic aortic rings. RESULTS: Treatment with IL-8/CXCL8 greatly increased 12-LO mRNA expression and protein production compared to treatment with angiotensin II. IL-8/CXCL8 also increased the expression of the AT1 receptor. The increase in 12-LO induced by IL-8/CXCL8 was inhibited by treatment with an AT1 receptor antagonist. The induction of 12-LO mRNA production and the proliferation of SHR VSMC by IL-8/CXCL8 was mediated by the ERK pathway. The proliferation of SHR VSMC and the vascular contraction in the thoracic aortic ring, both of which were induced by IL-8/CXCL8, were inhibited by baicalein, a 12-LO inhibitor. CONCLUSION: These results suggest that the potential role of IL-8/CXCL8 in hypertensive processes is likely mediated through the 12-LO pathway.
MeSH Terms
Figure
-

Figure 1 IL-8/CXCL8 increases 12-LO mRNA expression and protein production in SHR VSMC. (A) SHR and WKY VSMC were isolated from the thoracic aorta and cultured on plastic dishes at early passages (3 to 7). Total RNA was analyzed by real-time PCR. Bars represent means±SD from three independent experiments. *p<0.05 vs. WKY VSMC. (B) WKY VSMC were untreated or treated with IL-8/CXCL8 (100 ng/ml) for 2 h. Total RNA was then analyzed by real-time PCR. Bars represent means±SD from three independent experiments. (C) SHR VSMC were untreated or treated with IL-8/CXCL8 (100 ng/ml) or Ang II (100 nmol/L) for 2 h. Total RNA was then analyzed by real-time PCR. Bars represent means±SD from three independent experiments. a: p<0.05 vs. untreated VSMC. b: p<0.05 vs. treated with IL-8/CXCL8. (D) SHR VSMC were untreated or treated with IL-8/CXCL8 (100 ng/ml) or Ang II (100 nmol/L) for 2 and 4 h. Cell lysates were separated on 10% SDS-polyacrylamide gels and then immunoblotted with the 12-LO antibody. Data shown are representative of three independent experiments.

Figure 2 IL-8/CXCL8 increases AT1 receptor expression in SHR VSMC, and IL-8/CXCL8-induced expression of 12-LO mRNA is mediated through the AT1 receptor. (A) WKY and SHR VSMC were untreated or treated with IL-8/CXCL8 (100 ng/ml) for 2 h, and total RNA was analyzed by real-time PCR. Bars represent means±SD from four independent experiments. *p<0.05 vs. untreated VSMC. (B) VSMC were untreated or treated with IL-8/CXCL8 (100 ng/ml) for 2 and 4 h. Cell lysates were separated on 10% SDS-polyacrylamide gels and then immunoblotted with the AT1 receptor antibody. Data shown are representative of three independent experiments. (C) VSMC were untreated or treated with IL-8/CXCL8 (100 ng/ml) or IL-8/CXCL8 plus losartan (AT1 receptor antagonist, 10µmol/L) for 2 h or 4 h, and the total RNA and cell lysates were isolated. Total RNA was analyzed by real-time PCR. Cell lysates were separated on 10% SDS-polyacrylamide gels and then immunoblotted with the 12-LO antibody. Bars represent means±SD from three independent experiments. *p<0.05 vs. VSMC treated with IL-8/CXCL8. Data shown are representative of three independent experiments.
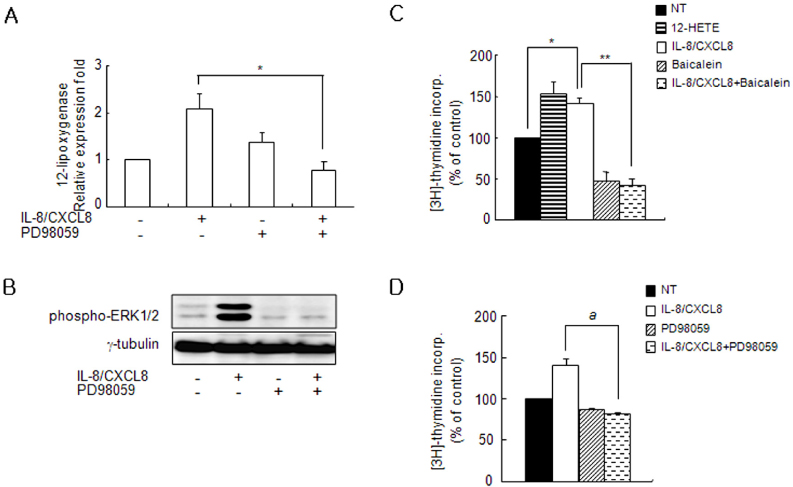
Figure 3 Expression of IL-8/CXCL8-induced 12-LO is mediated through the ERK pathway, and proliferation of SHR VSMC by IL-8/CXCL8 is inhibited by baicalein and PD98059. (A, B) VSMC were untreated (NT) or pretreated with PD98059 (ERK inhibitor, 10 µM) for 30 min. Cells were left untreated or treated with IL-8/CXCL8 (100 ng/ml) for 2 h, and the total RNA and cell lysates were isolated. The total RNA was analyzed by real-time PCR (A), and cell lysates were separated on 10% SDS-polyacrylamide gels and then immunoblotted with the phospho-ERK1/2 antibody (B). Bars represent means±SD from three independent experiments. *p<0.05 vs. VSMC treated with IL-8/CXCL8. Data shown are representative of three independent experiments. (C, D) SHR VSMC were treated with 12-HETE (500 nmol/L), with IL-8/CXCL8 (100 ng/ml), with baicalein (12-LO inhibitor, 10 µmol/L, B), or with PD98059 (10 µmol/L, C) for 48 h in medium containing [3H]-thymidine (1 µCi/ml). [3H]-thymidine incorporation is shown on the Y-axis. Bars represent means±SD from three independent experiments run in triplicate. *p<0.05 vs. untreated VSMC. **p<0.01 vs. VSMC treated with IL-8/CXCL8 alone. a: p<0.05 vs. VSMC treated with IL-8/CXCL8 alone.
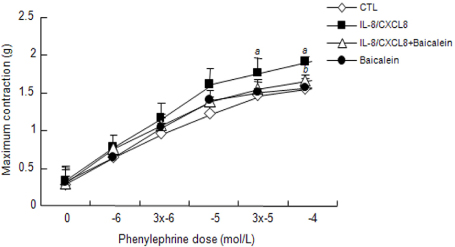
Figure 4 Effect of IL-8/CXCL8 on the phenylephrine-induced contraction of thoracic aortic rings. Phenylephrine (1~100 µmol/L) was added to isolated thoracic aortic rings pretreated with IL-8/CXCL8 (200 ng/mL) and/or the 12-LO inhibitor baicalein (10 µmol/L) for 1 h. All contractions are expressed as grams (g) of contractile tension in control rings not exposed to IL-8/CXCL8 or baicalein. Data are means±SD of three independent experiments. a: p<0.05 vs. control rings. b: p<0.05 vs. rings treated with IL-8/CXCL8 alone.
Cited by 1 articles
-
CCL5 Inhibits Elevation of Blood Pressure and Expression of Hypertensive Mediators in Developing Hypertension State Spontaneously Hypertensive Rats
Hye Young Kim, Hye Ju Cha, Jin Hee Choi, Young Jin Kang, So Young Park, Hee Sun Kim
J Bacteriol Virol. 2015;45(2):138-150. doi: 10.4167/jbv.2015.45.2.138.
Reference
-
1. Alexander RW. Hypertension and the pathogenesis of atherosclerosis. Oxidative stress and the mediation of arterial inflammatory response: a new perspective. Hypertension. 1995. 25:155–161.
Article2. Capers Q 4th, Alexander RW, Lou P, De Leon H, Wilcox JN, Ishizaka N, Howard AB, Taylor WR. Monocyte chemoattractant protein-1 expression in aortic tissues of hypertensive rats. Hypertension. 1997. 30:1397–1402.
Article3. Zhang Y, Griendling KK, Dikalova A, Owens GK, Talyor WR. Vascular hypertrophy in angiotensin II-induced hypertension is mediated by vascular smooth muscle cell-derived H2O2. Hypertension. 2005. 46:732–737.
Article4. Rodríguez-Iturbe B, Vaziri ND, Herrera-Acosta J, Johnson RJ. Oxidative stress, renal infiltration of immune cells and salt-sensitive hypertension: all for one and one for all. Am J Physiol Renal Physiol. 2004. 286:F606–F616.
Article5. Dhungana S, Sharrack B, Woodroofe N. Cytokines and chemokines in idiopathic intracranial hypertension. Headache. 2009. 49:282–285.
Article6. Gerszten RE, Garcia-Zepeda EA, Lim YC, Yoshida M, Ding MA, Gimbrone MA, Luster AD, Luscinskas FW, Rosenzweig A. MCP-1 and IL-8 trigger firm adhesion of monocytes to vascular endothelium under flow conditions. Nature. 1999. 398:718–723.
Article7. Boekholdt S, Peters R, Hack CE, Day NE, Luben R, Bingham SA, Wareham NJ, Reitsma PH, Khaw KT. IL-8 plasma concentrations and the risk of future coronary artery disease in apparently healthy men and women. Arterioscler Thromb Vasc Biol. 2004. 24:1503–1508.
Article8. Kim HY, Kang YJ, Song IH, Choi HC, Kim HS. Upregulation of IL-8/CXCL8 in vascular smooth muscle cells from spontaneously hypertensive rats. Hypertens Res. 2008. 31:515–523.
Article9. Natarajan R, Rosdahl J, Gonzales N, Bai W. Regulation of 12-lipoxygenase by cytokines in vascular smooth muscle cells. Hypertension. 1997. 30:873–879.
Article10. Preston IR, Hill NS, Warburton RR, Fanburg BL. Role of 12-lipoxygenase in hypoxia-induced rat pulmonary artery smooth muscle cell proliferation. Am J Physiol Lung Cell Mol Physiol. 2006. 290:L367–L374.
Article11. Merched AJ, Ko K, Gotlinger KH, Serhan CN, Chan L. Atherosclerosis: evidence for impairment of resolution of vascular inflammation governed by specific lipid mediators. FASEB J. 2008. 22:3595–3606.
Article12. Reddy MA, Sahar S, Villeneuve LM, Lanting L, Natarajan R. Role of Src tyrosine kinase in the atherogenic effects of the 12/15-lipoxygenase pathway in vascular smooth muscle cells. Arterioscler Thromb Vasc Biol. 2009. 29:387–393.
Article13. Natarajan R, Gu JL, Rossi J, Gonzales N, Lanting L, Xu L, Nadler J. Elevated glucose angiontensin II increase 12-lipoxygenase activity and expression in porcine aortic smooth muscle cells. Proc Natl Acad Sci USA. 1993. 90:4947–4951.
Article14. Kim JA, Gu JL, Natarajan R, Rerliner JA, Nadler JL. A leukocyte type 12-lipoxygenase is expressed in human vascular and mononuclear cells: Evidence for upregulation by angiotensin II. Arterioscler Thromb Vasc Biol. 1995. 15:942–948.
Article15. Sasaki M, Hori MT, Hino T, Golub MS, Tuck ML. Elevated 12-lipoxygenase activity in the spontaneously hypertensive rat. Am J Hypertens. 1997. 10:371–378.
Article16. Yue TL, Wang X, Sung CP. Interleukin-8: a mitogen and chemoattractant for vascular smooth muscle cells. Circ Res. 1994. 75:1–7.
Article17. Usui M, Egashira K, Tomita H, Koyanagi M, Katoh M, Shimokawa H, Takeya M, Yoshimura T, Matsushima K, Takeshita A. Important role of local angiotensin II activity mediated via type 1 receptor in the pathogenesis of cardiovascular inflammatory changes induced by chronic blockade of nitric oxide synthesis in rats. Circulation. 2000. 101:305–310.
Article18. Wolf G, Wenzel U, Burns KD, Harris RC, Stahl AK, Thaiss F. Angiotensin II activates nuclear transcription factor-κB through AT1 and AT2 receptors. Kidney Int. 2002. 61:1986–1995.
Article19. Siragy HM. AT1 and AT2 receptors in the kidney: Role of disease and treatment. Am J Kidney Dis. 2000. 36:Suppl 1. S4–S9.20. Horiuchi M, Akishita M, Dzau VJ. Recent progress in angiotensin II Type 2 receptor research in the cardiovascular system. Hypertension. 1999. 33:613–621.
Article21. Jin XQ, Fukuda N, Lai YM, Suzuki R, Tahira Y, Takgi H, Ikeda Y, Kanmatsuse K, Miyazaki H. Angiotensin II type 2 receptor gene transfer downregulates angiotensin II type 1a receptor in vascular smooth muscle cells. Hypertension. 2002. 39:1021–1027.
Article22. Reddy MA, Adler SG, Kim YS, Lanting L, Rossi J, Kang SW, Nadler JL, Shahed A, Natarajan R. Interaction of MAPK and 12-lipoxygenase pathways in growth and matrix protein expression in mesangial cells. Am J Physiol Renal Physiol. 2002. 283:F985–F994.23. Buemi M, Marino D, Floccari F, Ruello A, Nostro L, Aloisi C, Marino MT, Pasquale G, Corica F. Effect of interleukin 8 and ICAM-1 on calcium-dependent outflow of K+ in erythrocytes from subjects with essential hypertension. Curr Med Res Opin. 2004. 20:19–24.
Article24. Ohkawa F, Ikeda U, Kanbe T, Kawasaki K, Shimada K. Effects of inflammatory cytokines on vascular tone. Cardiovasc Res. 1995. 30:711–715.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Expression of Endothelin-1 by Stimulation with CXCL8 in Mouse Peritoneal Macrophages
- 15-Deoxy-Delta(12,14)-Prostaglandin J2 Upregulates the Expression of LPS-Induced IL-8/CXCL8 mRNA in Vascular Smooth Muscle Cells from Spontaneously Hypertensive Rats
- Downregulation of Angiotensin II-Induced 12-Lipoxygenase Expression and Cell Proliferation in Vascular Smooth Muscle Cells from Spontaneously Hypertensive Rats by CCL5
- Sulfatase 1 mediates the inhibitory effect of angiotensin II type 2 receptor inhibitor on angiotensin II-induced hypertensive mediator expression and proliferation in vascular smooth muscle cells from spontaneously hypertensive rats
- CCL5 Inhibits Elevation of Blood Pressure and Expression of Hypertensive Mediators in Developing Hypertension State Spontaneously Hypertensive Rats
