Disruption of Microtubules Sensitizes the DNA Damage-induced Apoptosis Through Inhibiting Nuclear Factor kappaB (NF-kappaB) DNA-binding Activity
- Affiliations
-
- 1Department of Pharmacology, Research Institute for Medical Science, Daejeon Regional Cancer Center, Daejeon, Korea. gmhur@cnu.ac.kr
- 2Division of Rheumatology, Department of Internal Medicine, College of Medicine, Chungnam National University, Daejeon, Korea.
- KMID: 1779226
- DOI: http://doi.org/10.3346/jkms.2010.25.11.1574
Abstract
- The massive reorganization of microtubule network involves in transcriptional regulation of several genes by controlling transcriptional factor, nuclear factor-kappa B (NF-kappaB) activity. The exact molecular mechanism by which microtubule rearrangement leads to NF-kappaB activation largely remains to be identified. However microtubule disrupting agents may possibly act in synergy or antagonism against apoptotic cell death in response to conventional chemotherapy targeting DNA damage such as adriamycin or comptothecin in cancer cells. Interestingly pretreatment of microtubule disrupting agents (colchicine, vinblastine and nocodazole) was observed to lead to paradoxical suppression of DNA damage-induced NF-kappaB binding activity, even though these could enhance NF-kappaB signaling in the absence of other stimuli. Moreover this suppressed NF-kappaB binding activity subsequently resulted in synergic apoptotic response, as evident by the combination with Adr and low doses of microtubule disrupting agents was able to potentiate the cytotoxic action through caspase-dependent pathway. Taken together, these results suggested that inhibition of microtubule network chemosensitizes the cancer cells to die by apoptosis through suppressing NF-kappaB DNA binding activity. Therefore, our study provided a possible anti-cancer mechanism of microtubule disrupting agent to overcome resistance against to chemotherapy such as DNA damaging agent.
Keyword
MeSH Terms
-
Animals
Antibiotics, Antineoplastic/therapeutic use
*Apoptosis
Caspases/metabolism
Cell Line
Colchicine/pharmacology
DNA/metabolism
*DNA Damage
Doxorubicin/therapeutic use
Humans
Mice
Microtubules/chemistry/*drug effects/metabolism
NF-kappa B/antagonists & inhibitors/*metabolism
Neoplasms/drug therapy
Nocodazole/pharmacology
Protein Binding
Signal Transduction
Tubulin Modulators/*pharmacology
Vinblastine/pharmacology
Figure
-
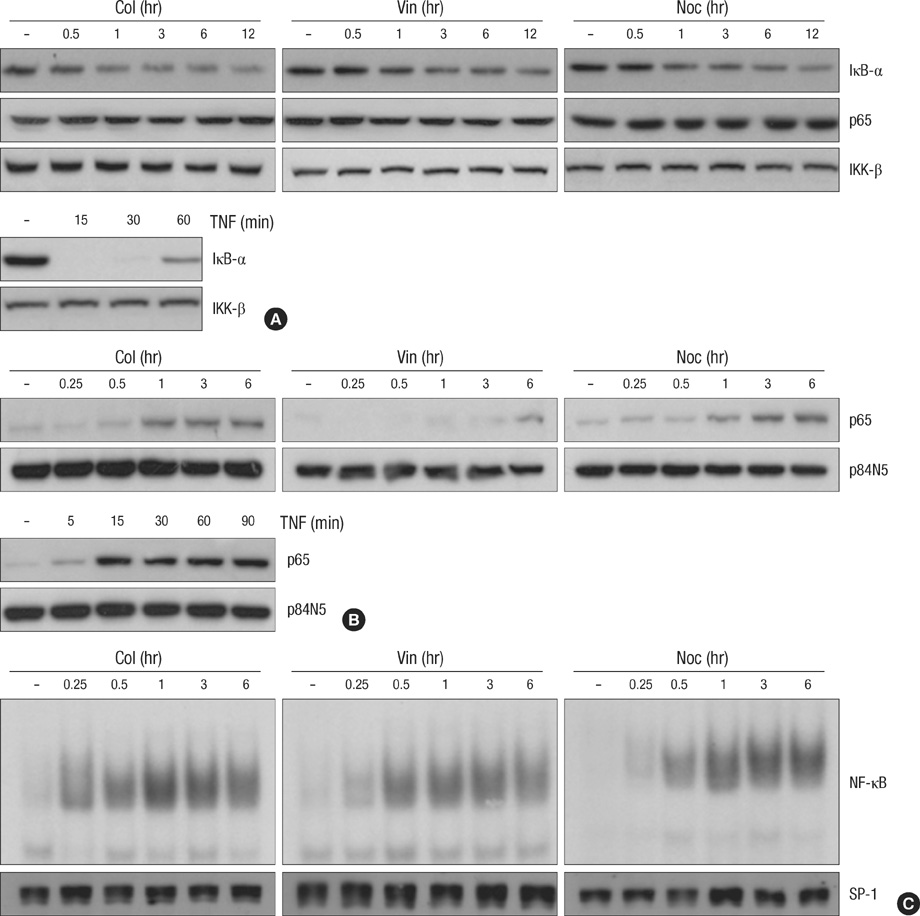
Fig. 1 Microtubule disrupting agents induces NF-κB activation through nuclear translocation of NF-κB subunit p65. (A) HeLa cells were treated with microtubule disrupting agents (upper panel) including colchicines (Col, 10 µM), vinblastine (Vin, 1 µM) and nocodazole (Noc, 0.5 µM) or TNF (lower panel) for various times as indicated. Cell extracts were applied to SDS-PAGE for immunoblotting with anti-IκB-α and anti-p65 antibodies. As a protein loading control, the same amounts of extracts were analyzes by immunoblotting with anti-IKK-β antibody. (B, C) After HeLa cells were treated with 10 µM of Col, 1 µM of Vin or 0.5 µM of Noc for various times as indicated, nuclear extracts were obtained as described under Materials and Methods. (B) The nuclear translocation of NF-κB subunit p65 was analyzed by immunoblotting with anti-p65 and anti-p84N5 antibodies from nuclear extracts from each sample. (C) EMSAs were performed with labeled NF-κB oligomers and 5 µg of nuclear extracts to analyze NF-κB binding activity. As a control, 50 µg of the nuclear extracts were applied to SDS-PAGE for immunoblotting with anti-SP1 antibody.
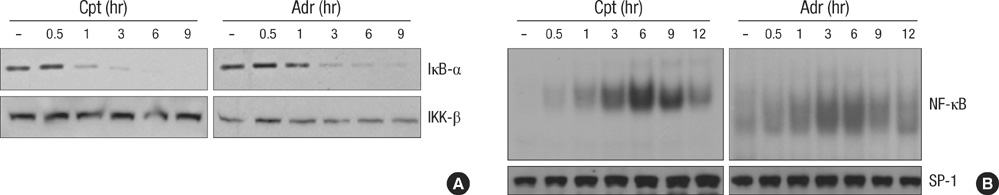
Fig. 2 DNA damaging agents induces NF-κB activation. (A) HeLa cells were treated with adriamycin (Adr, 10 µg/mL) or comptothecin (Cpt, 100 µM) for various times as indicated. Cell extracts were applied to SDS-PAGE for immunoblotting with anti-IκB-α and anti-IKK-β antibodies. (B) HeLa cells were treated as described in A for various times as indicated. Nuclear extracts were prepared and 5 µg of nuclear extracts from each sample was used to analyze NF-κB binding activity by EMSA with a NF-κB probe.
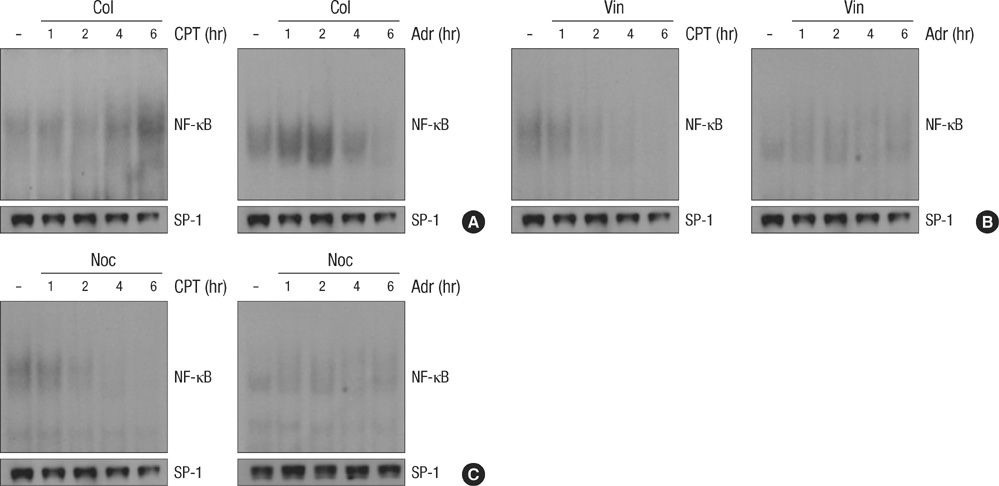
Fig. 3 Microtubule disrupting agents suppresses the DNA damage-induced NF-κB binding activity. HeLa cells were treated with Adr (10 µg/mL) or Cpt (100 µM) for various times as indicated in the presence or absence of Col (A, 10 µM), Vin (B, 1 µM) and Noc (C, 0.5 µM). Nuclear extracts were prepared and 5 µg of nuclear extracts from each sample was used to analyze NF-κB binding activity by EMSA as described in Fig. 2B. As a control, 50 µg of the nuclear extracts were applied to SDS-PAGE for immunoblotting with anti-SP1 antibody.
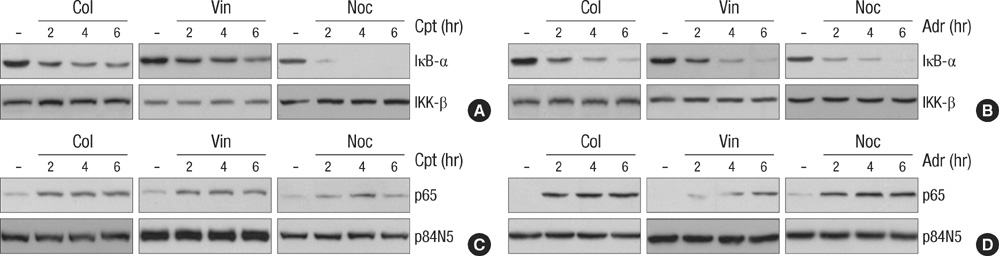
Fig. 4 Microtubule disrupting agents does not affect the DNA damage-induced IκB-α degradation and nuclear translocation of NF-κB subunit p65. HeLa cells were treated with Adr (10 µg/mL) or Cpt (100 µM) for various times as indicated in the presence or absence of Col (10 µM), Vin (1 µM) and Noc (0.5 µM). (A, B) Total cell extracts were prepared and applied to SDS-PAGE for immunoblotting with anti-IκB-α and anti-IKK-β antibodies. (C, D) Nuclear extracts were obtained as described Fig.1B, and the nuclear translocation of NF-κB subunit p65 was analyzed by immunoblotting with anti-p65 and anti-p84N5 antibodies from nuclear extracts (50 µg) from each sample.

Fig. 5 Microtubule disrupting agents sensitizes DNA damage-induced apoptotic cell death. (A) HeLa cells were treated with Adr (10 µg/mL) for various times as indicated in the presence or absence of pan-caspase inhibitor z-VAD-FMK (20 µM) and cell extracts were analyzed by immunoblotting with antibodies against casapse-9, -3 and PARP. As a protein loading control, the same amounts of extracts were analyzes by immunoblotting with anti-actin antibody. The asterisks indicate cleaved products of casapse-3 and PARP upon Adr treatment. (B) Wild-type and p65-/- MEF cells were treated with various concentrations of Adr as indicated. 12 hr after treatment, the percentage of cell death was determined by trypan blue exclusion assay as described under Materials and Methods. The results represent the mean values of at least three independent experiments. *, P<0.05, compared with Adr-treated wild-type MEF cells. (C) The expression levels of p65 and IKK-β in wild-type and p65-/- MEF cells. The equal amount of cell extract from each cells was analyzed by immunoblotting with antibodies against p65, IKK-β and actin. (D) HeLa cells were pretreated with the NF-κB specific inhibitors, 5 µM Bay-11 or 1 µM TPCA-1 for 30 min and then treated with 10 µg/mL Adr for 12 hr. Cell death was quantified as described in (B), and each column shows mean±S.E. of at least three independent experiments. *, P<0.05, compared with Adr-treated group. (E) HeLa cells were treated with Adr (10 µg/mL) for 15 hr in the presence or absence of Col (10 µM), Vin (1 µM) and Noc (0.5 µM), or z-VAD-FMK (20 µM). Then, cells were visualized with a normal light microscope with an inverted microscope. (F) HeLa cells were treated with Adr (10 µg/mL) for various times as indicated in the presence or absence of Col (10 µM), Vin (1 µM) and Noc (0.5 µM). Cell death was quantified as described in (B), and each column shows mean±S.E. of at least three independent experiments. (G) HeLa cells were treated as described in (C). Cell extracts form each sample were analyzed by SDS-PAGE followed by immunoblotting with antibodies against casapse-9, -3, PARP and actin. The asterisks indicate cleaved products of casapse-3 and PARP.
Reference
-
1. Karin M, Greten FR. NF-kappaB: linking inflammation and immunity to cancer development and progression. Nat Rev Immunol. 2005. 5:749–759.2. Lin Y, Bai L, Chen W, Xu S. The NF-kappaB activation pathways, emerging molecular targets for cancer prevention and therapy. Expert Opin Ther Targets. 2010. 14:45–55.3. Brown K, Gerstberger S, Carlson L, Franzoso G, Siebenlist U. Control of I kappa B-alpha proteolysis by site-specific, signal-induced phosphorylation. Science. 1995. 267:1485–1488.
Article4. Hur GM, Lewis J, Yang Q, Lin Y, Nakano H, Nedospasov S, Liu ZG. The death domain kinase RIP has an essential role in DNA damage-induced NF-kappaB activation. Genes Dev. 2003. 17:873–882.5. Waddick KG, Uckun FM. Innovative treatment programs against cancer: II. Nuclear factor-kappaB (NF-kappaB) as a molecular target. Biochem Pharmacol. 1999. 57:9–17.6. Daroczi B, Kari G, Ren Q, Dicker AP, Rodeck U. Nuclear factor kappaB inhibitors alleviate and the proteasome inhibitor PS-341 exacerbates radiation toxicity in zebrafish embryos. Mol Cancer Ther. 2009. 8:2625–2634.7. Li F, Sethi G. Targeting transcription factor NF-kappaB to overcome chemoresistance and radioresistance in cancer therapy. Biochim Biophys Acta. 2010. 1805:167–180.8. Egan LJ, Eckmann L, Greten FR, Chae S, Li ZW, Myhre GM, Robine S, Karin M, Kagnoff MF. IkappaB-kinasebeta-dependent NF-kappaB activation provides radioprotection to the intestinal epithelium. Proc Natl Acad Sci USA. 2004. 101:2452–2457.9. Starenki D, Namba H, Saenko V, Ohtsuru A, Yamashita S. Inhibition of nuclear factor-kappaB cascade potentiates the effect of a combination treatment of anaplastic thyroid cancer cells. J Clin Endocrinol Metab. 2004. 89:410–418.10. Ahmed KM, Li JJ. ATM-NF-kappaB connection as a target for tumor radiosensitization. Curr Cancer Drug Targets. 2007. 7:335–342.11. Fan Y, Dutta J, Gupta N, Fan G, Gélinas C. Regulation of programmed cell death by NF-kappaB and its role in tumorigenesis and therapy. Adv Exp Med Biol. 2008. 615:223–250.12. Nadiminty N, Lou W, Sun M, Chen J, Yue J, Kung HJ, Evans CP, Zhou Q, Gao AC. Aberrant activation of the androgen receptor by NF-kappaB2/p52 in prostate cancer cells. Cancer Res. 2010. 70:3309–3319.13. Puvvada SD, Funkhouser WK, Greene K, Deal A, Chu H, Baldwin AS, Tepper JE, O'Neil BH. NF-kB and Bcl-3 activation are prognostic in metastatic colorectal cancer. Oncology. 2010. 78:181–188.14. Rosette C, Karin M. Cytoskeletal control of gene expression: depolymerization of microtubules activates NF-kappaB. J Cell Biol. 1995. 128:1111–1119.15. Das KC, White CW. Activation of NF-kappaB by antineoplastic agents. Role of protein kinase C. J Biol Chem. 1997. 272:14914–14920.16. Mackenzie GG, Keen CL, Oteiza PI. Microtubules are required for NF-kappaB nuclear translocation in neuroblastoma IMR-32 cells: modulation by zinc. J Neurochem. 2006. 99:402–415.17. Mikenberg I, Widera D, Kaus A, Kaltschmidt B, Kaltschmidt C. Transcription factor NF-kappaB is transported to the nucleus via cytoplasmic dynein/dynactin motor complex in hippocampal neurons. PLoS One. 2007. 2:e589.18. Devary Y, Rosette C, DiDonato JA, Karin M. NF-kappaB activation by ultraviolet light not dependent on a nuclear signal. Science. 1993. 261:1442–1445.19. Doostzadeh-Cizeron J, Terry NH, Goodrich DW. The nuclear death domain protein p84N5 activates a G2/M cell cycle checkpoint prior to the onset of apoptosis. J Biol Chem. 2001. 276:1127–1132.
Article20. Kim R, Emi M, Tanabe K. Caspase-dependent and -independent cell death pathways after DNA damage (Review). Oncol Rep. 2005. 14:595–599.
Article21. Berenson JR, Ma HM, Vescio R. The role of nuclear factor-kappaB in the biology and treatment of multiple myeloma. Semin Oncol. 2001. 28:626–633.22. Um JH, Kang CD, Lee BG, Kim DW, Chung BS, Kim SH. Increased and correlated nuclear factor-kappa B and Ku autoantigen activities are associated with development of multidrug resistance. Oncogene. 2001. 20:6048–6056.
Article23. Meulmeester E, Jochemsen AG. p53: a guide to apoptosis. Curr Cancer Drug Targets. 2008. 8:87–97.
Article24. Green DR, Kroemer G. Cytoplasmic functions of the tumour suppressor p53. Nature. 2009. 458:1127–1130.
Article25. Schuler M, Green DR. Mechanisms of p53-dependent apoptosis. Biochem Soc Trans. 2001. 29:684–688.
Article26. Shen Y, White E. p53-dependent apoptosis pathways. Adv Cancer Res. 2001. 82:55–84.
Article27. el-Deiry WS. Role of oncogenes in resistance and killing by cancer therapeutic agents. Curr Opin Oncol. 1997. 9:79–87.
Article28. Pérez-Tomás R. Multidrug resistance: retrospect and prospects in anti-cancer drug treatment. Curr Med Chem. 2006. 13:1859–1876.29. Németh ZH, Deitch EA, Davidson MT, Szabó C, Vizi ES, Haskó G. Disruption of the actin cytoskeleton results in nuclear factor-kappaB activation and inflammatory mediator production in cultured human intestinal epithelial cells. J Cell Physiol. 2004. 200:71–81.30. Mikenberg I, Widera D, Kaus A, Kaltschmidt B, Kaltschmidt C. TNF-alpha mediated transport of NF-kappaB to the nucleus is independent of the cytoskeleton-based transport system in non-neuronal cells. Eur J Cell Biol. 2006. 85:529–536.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Differential Regulation of NF-kappaB Signaling during Human Cytomegalovirus Infection
- Rifampicin Inhibits the LPS-induced Expression of Toll-like Receptor 2 via the Suppression of NF-kappaB DNA-binding Activity in RAW 264.7 Cells
- The Proteasome Inhibitor MG132 Sensitizes Lung Cancer Cells to TRAIL-induced Apoptosis by Inhibiting NF-kappaB Activation
- beta-Lapachone suppresses radiation-induced activation of nuclear factor-kappaB
- The Inhibitory Effect of Epicatechin on IL-1beta -induced iNOS Expression and NO Production in RINm5F Cell
