Requirement of Pretone by Thromboxane A2 for Hypoxic Pulmonary Vasoconstriction in Precision-cut Lung Slices of Rat
- Affiliations
-
- 1Department of Physiology, Seoul National University College of Medicine, Seoul 110-799, Korea. sjoonkim@snu.ac.kr
- 2Ischemic/Hypoxic Disease Institute, Seoul National University, Seoul 110-799, Korea.
- 3Kidney Institute Medical Research Center, Seoul National University, Seoul 110-799, Korea.
- 4Department of Anesthesiology and Pain Medicine, Samsung Medical Center, Sungkyunkwan University School of Medicine, Seoul 135-710, Korea.
- KMID: 2285441
- DOI: http://doi.org/10.4196/kjpp.2012.16.1.59
Abstract
- Hypoxic pulmonary vasoconstriction (HPV) is physiologically important response for preventing mismatching between ventilation and perfusion in lungs. The HPV of isolated pulmonary arteries (HPV-PA) usually require a partial pretone by thromboxane agonist (U46619). Because the HPV of ventilated/perfused lungs (HPV-lung) can be triggered without pretone conditioning, we suspected that a putative tissue factor might be responsible for the pretone of HPV. Here we investigated whether HPV can be also observed in precision-cut lung slices (PCLS) from rats. The HPV in PCLS also required partial contraction by U46619. In addition, K+ channel blockers (4AP and TEA) required U46619-pretone to induce significant contraction of PA in PCLS. In contrast, the airways in PCLS showed reversible contraction in response to the K+ channel blockers without pretone conditioning. Also, the airways showed no hypoxic constriction but a relaxation under the partial pretone by U46619. The airways in PCLS showed reliable, concentration-dependent contraction by metacholine (EC50, ~210 nM). In summary, the HPV in PCLS is more similar to isolated PA than V/P lungs. The metacholine-induced constriction of bronchioles suggested that the PLCS might be also useful for studying airway physiology in situ.
MeSH Terms
Figure
-
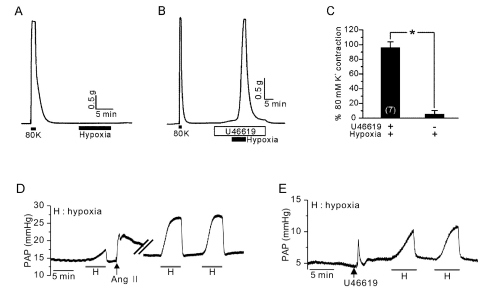
Fig. 1 Essential role of TXA2 on HPV in PAs. (A) Hypoxia (3% Po2) alone did not induce an increased tension of PAs. (B) In the presence of U46619 (10 nM), a strong contraction of PAs occurred in response to hypoxia. (C) Summaries of PAs tone normalized to 80K-induced contraction were shown as a bar graph (mean±SEM, n=7). *Indicates statistically significant difference from the control value (p<0.05). (D) Induction of HPV in V/P lungs. The increase in pulmonary arterial pressure (PAP, mmHg) was observed by hypoxic ventilation. In the same lung, Ang II induced a transient PAP increase, which augmented the HPV response. (E) A pretreatment with U46619 also induced a transient PAP increase, and HPV was also observed afterwards.
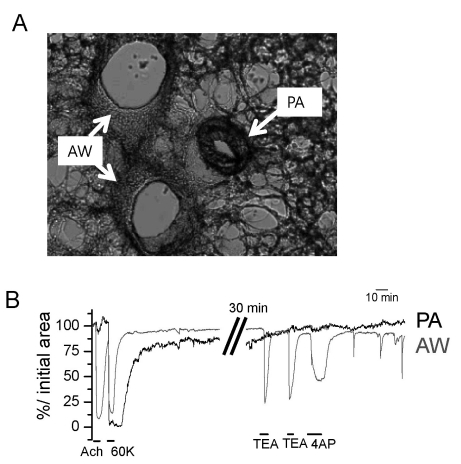
Fig. 2 Identification of PA and AW constrictions in rat PCLS. (A) A representative image of rat PCLS demonstrating the AW and PA. (B) Functional discrimination of PA and AW according to differential responses to ACh and K+ channel blockers. AW showed strong constriction to ACh (10 µM), tetraethylammonium (TEA, 2 mM) and 4-aminopyridine (4AP, 5 mM) while PA showed no response to the concomitant application of the above agents.
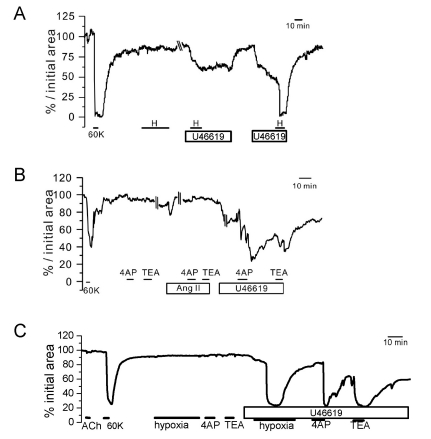
Fig. 3 Responses of PA to hypoxia and K+ channel blockers under pretone conditions in rat PCLS. Three representative cases of relative luminal area (% initial area) changes in PAs. In all cases (A~C), the K+ channel blockers (TEA and 4AP) induced reversible constriction only under the pretreatment with 5 nM U46619 while not with 100 nM Ang II (B). Also, the hypoxic constriction was observed under the pretreatment with 5 nM U46619 (A, C).
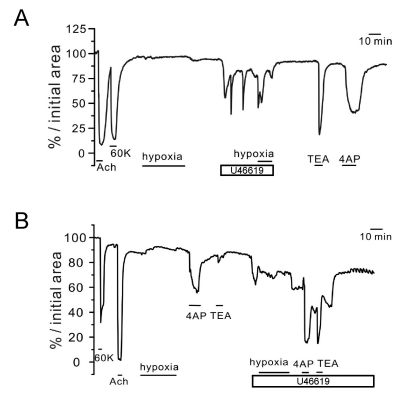
Fig. 4 Responses of AW to hypoxia and K+ channel blockers under pretone conditions in rat PCLS. Two representative cases of luminal area (% initial area) changes in AWs. The strong contractile response to 10 µM ACh was confirmed along with the response to 60 K. Hypoxia alone had no effect (A) or induced a slight dilation (B) of AW. Application of 5 nM U46619 induced oscillatory or tonic constrictions that were not affected (A) or partly inhibited by hypoxia (B). Different from PAs, both 4AP and TEA induced constriction of AW in the absence of U46619 pretreatment.
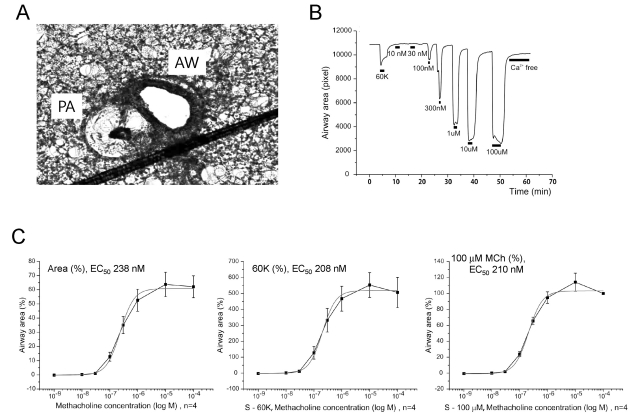
Fig. 5 Response of AW to metacholine (MCh) in mouse PCLS. (A) A representative image of rat PCLS demonstrating the AW and PA. Note the relatively narrow lumen of PA in the mouse PCLS. (B) Concentration-dependent reversible constrictions of mouse AW. The maximum constriction induced by 100 µM MCh was markedly larger (>five fold) than the 60K-constriction. (C) Normalized AW constrictions to different concentrations of MCh were averaged and fitted to logistic functions. Each panel was obtained by different type of normalization criteria; relative change from the maximum luminal area (left), normalized against 60K constriction (middle), and normalized to maximum constriction by 100 µM MCh (right). Regardless of the normalization criteria, the half-effective concentration (EC50) of MCh located at similar ranges.
Reference
-
1. Bonnet S, Dubuis E, Vandier C, Martin S, Marthan R, Savineau JP. Reversal of chronic hypoxia-induced alterations in pulmonary artery smooth muscle electromechanical coupling upon air breathing. Cardiovasc Res. 2002; 53:1019–1028. PMID: 11922912.
Article2. Osipenko ON, Alexander D, MacLean MR, Gurney AM. Influence of chronic hypoxia on the contributions of non-inactivating and delayed rectifier K currents to the resting potential and tone of rat pulmonary artery smooth muscle. Br J Pharmacol. 1998; 124:1335–1337. PMID: 9723941.
Article3. Yoo HY, Park SJ, Seo EY, Park KS, Han JA, Kim KS, Shin DH, Earm YE, Zhang YH, Kim SJ. Role of thromboxane A2-activated nonselective cation channels in hypoxic pulmonary vasoconstriction of rat. Am J Physiol Cell Physiol. 2012; 302:C307–C317. PMID: 21998141.
Article4. Leach RM, Robertson TP, Twort CH, Ward JP. Hypoxic vasoconstriction in rat pulmonary and mesenteric arteries. Am J Physiol. 1994; 266:L223–L231. PMID: 8166292.
Article5. Rodman DM, Yamaguchi T, O'Brien RF, McMurtry IF. Hypoxic contraction of isolated rat pulmonary artery. J Pharmacol Exp Ther. 1989; 248:952–959. PMID: 2467984.6. Park SJ, Yoo HY, Earm YE, Kim SJ, Kim JK, Kim SD. Role of arachidonic acid-derived metabolites in the control of pulmonary arterial pressure and hypoxic pulmonary vasoconstriction in rats. Br J Anaesth. 2011; 106:31–37. PMID: 20935003.
Article7. Sanderson MJ. Exploring lung physiology in health and disease with lung slices. Pulm Pharmacol Ther. 2011; 24:452–465. PMID: 21600999.
Article8. Struckmann N, Schwering S, Wiegand S, Gschnell A, Yamada M, Kummer W, Wess J, Haberberger RV. Role of muscarinic receptor subtypes in the constriction of peripheral airways: studies on receptor-deficient mice. Mol Pharmacol. 2003; 64:1444–1451. PMID: 14645675.
Article9. Moreno L, Perez-Vizcaino F, Harrington L, Faro R, Sturton G, Barnes PJ, Mitchell JA. Pharmacology of airways and vessels in lung slices in situ: role of endogenous dilator hormones. Respir Res. 2006; 7:111. PMID: 16923180.
Article10. Paddenberg R, König P, Faulhammer P, Goldenberg A, Pfeil U, Kummer W. Hypoxic vasoconstriction of partial muscular intra-acinar pulmonary arteries in murine precision cut lung slices. Respir Res. 2006; 7:93. PMID: 16808843.
Article11. Bai Y, Sanderson MJ. Modulation of the Ca2+ sensitivity of airway smooth muscle cells in murine lung slices. Am J Physiol Lung Cell Mol Physiol. 2006; 291:L208–L221. PMID: 16461427.12. Martin C, Uhlig S, Ullrich V. Cytokine-induced bronchoconstriction in precision-cut lung slices is dependent upon cyclooxygenase-2 and thromboxane receptor activation. Am J Respir Cell Mol Biol. 2001; 24:139–145. PMID: 11159047.
Article13. Bergner A, Sanderson MJ. Acetylcholine-induced calcium signaling and contraction of airway smooth muscle cells in lung slices. J Gen Physiol. 2002; 119:187–198. PMID: 11815668.
Article14. Brueggemann LI, Kakad PP, Love RB, Solway J, Dowell ML, Cribbs LL, Byron KL. Kv7 potassium channels in airway smooth muscle cells: signal transduction intermediates and pharmacological targets for bronchodilator therapy. Am J Physiol Lung Cell Mol Physiol. 2012; 302:L120–L132. PMID: 21964407.
Article15. Ressmeyer AR, Larsson AK, Vollmer E, Dahlèn SE, Uhlig S, Martin C. Characterisation of guinea pig precision-cut lung slices: comparison with human tissues. Eur Respir J. 2006; 28:603–611. PMID: 16737991.
Article16. Desireddi JR, Farrow KN, Marks JD, Waypa GB, Schumacker PT. Hypoxia increases ROS signaling and cytosolic Ca2+ in pulmonary artery smooth muscle cells of mouse lungs slices. Antioxid Redox Signal. 2010; 12:595–602. PMID: 19747064.17. Held HD, Martin C, Uhlig S. Characterization of airway and vascular responses in murine lungs. Br J Pharmacol. 1999; 126:1191–1199. PMID: 10205008.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Disappearance of Hypoxic Pulmonary Vasoconstriction and O2-Sensitive Nonselective Cationic Current in Arterial Myocytes of Rats Under Ambient Hypoxia
- The Effect of Isoproterenol, Dobutamine, and Milrinone on Pulmonary Vasoconstriction in Isolated Rat Lungs
- Effects of Glibenclamide and L-NAME on Hypoxic Pulmonary Vasoconstriction in Rats
- Pulmonary Hypertension Associated with Acute Hypoxic Pulmonary Vasoconstriction in a Patient with Acute Myeloid Leukemia
- PGF2 alpha causes bronchoconstriction and pulmonary vasoconstriction via thromboxane receptors in rat lung
