Bisphenol A and 4-tert-Octylphenol Inhibit Cx46 Hemichannel Currents
- Affiliations
-
- 1Department of Physiology, College of Medicine, Dankook University, Cheonan 330-714, Korea. seung@dku.edu
- KMID: 2071822
- DOI: http://doi.org/10.4196/kjpp.2015.19.1.73
Abstract
- Connexins (Cx) are membrane proteins and monomers for forming gap junction (GJ) channels. Cx46 and Cx50 are also known to function as conductive hemichannels. As part of an ongoing effort to find GJ-specific blocker(s), endocrine disruptors were used to examine their effect on Cx46 hemichannels expressed in Xenopus oocytes. Voltage-dependent gating of Cx46 hemichannels was characterized by slowly activating outward currents and relatively fast inward tail currents. Bisphenol A (BPA, 10 nM) reduced outward currents of Cx46 hemichannels up to ~18% of control, and its effect was reversible (n=5). 4-tert-Octylphenol (OP, 1 microM) reversibly reduced outward hemichannel currents up to ~28% (n=4). However, overall shapes of Cx46 hemichannel current traces (outward and inward currents) were not changed by these drugs. These results suggest that BPA and OP are likely to occupy the pore of Cx46 hemichannels and thus obstruct the ionic fluxes. This finding provides that BPA and OP are potential candidates for GJ channel blockers.
MeSH Terms
Figure
-
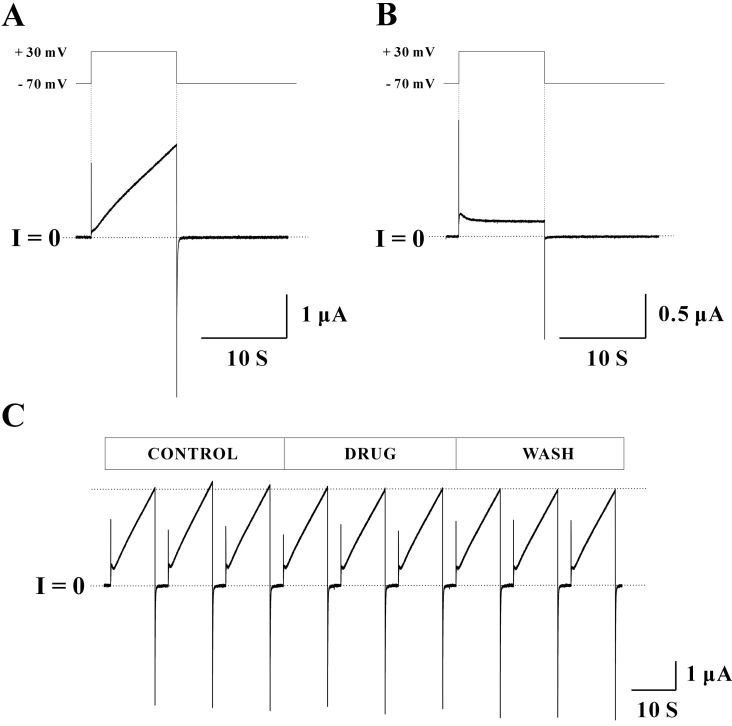
Fig. 1 Voltage-dependent gating of Cx46 hemichannel. (A) When holding the oocyte expressing Cx46 hemichannels at -70 mV, currents were near zero level. When stepping to a depolarizing potential at +30 mV for 10 sec, slowly activating outward currents were observed. When stepping back to the initial holding potential at -70 mV, relatively fast inward tail currents were shown. Immediately after, currents were back to zero level. The total recording time was 40 sec. (B) Same pulse protocol used in Fig. 1A was applied to un-injected oocyte. Slowly activating outward and relatively fast inward currents as those of Cx46 cRNA injected oocytes were not observed from the un-injected oocyte. (C) Same pulse protocol used in Fig. 1A was applied to each sweep. One set of records included a total of nine sweeps (3 of each control, drug, and wash phases, respectively) and were obtained from a single oocyte expressing Cx46 hemichannels. Each sweep record was acquired immediately after the perfusion with new bath solution. Most peak currents were quite stable for the entire recording period indicated by a dotted line. The voltage-dependence of Cx46 hemichannels represented by slowly activating outward currents and relatively fast inward currents were observed without any change. Current traces for about 25 seconds before the next sweep were omitted for data illustration.
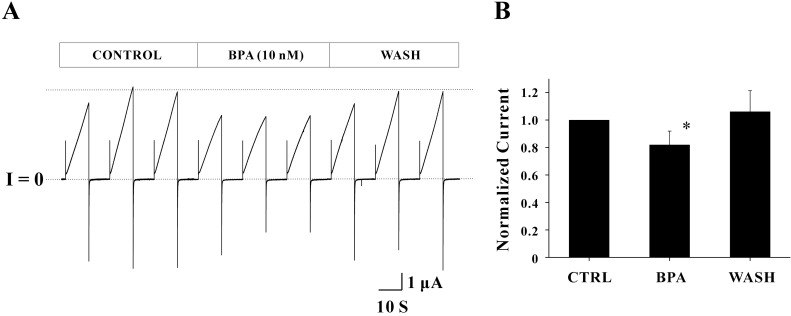
Fig. 2 The effect of BPA (10 nM) on Cx46 hemichannel currents. (A) One set of records obtained from a single oocyte expressing Cx46 hemichannels is shown. The pulse protocol included VH=-70 mV, Vstep=+30 mV for 10 sec, and a total recording time of 40 sec was used to each sweep. In the control phase (initial three sweeps), the average peak current was 4.75±0.48 µA. The average peak current after applying 10 nM BPA (middle three sweeps) was 3.45±0.05 µA, indicating that BPA inhibited Cx46 hemichannel currents to 27.36% of control. Reversible effects of BPA were observed. In the wash phase (last three sweeps) these reduced currents were returned to control level (4.64±0.40 µA). Overall shapes for outward and inward currents were not affected by BPA. Current traces for about 20 seconds before the next sweep were omitted for better illustration. (B) Normalized peak currents of each phase determined by the ratio of average peak currents of the drug (or wash) phase to those of the control phase are shown. Normalized peak currents with BPA treatment were 0.82±0.10 (n=5). This inhibitory effect of BPA (~18% reduction) was statistically significant (*p<0.05). However, normalized peak currents in the wash phase (1.06±0.15, n=5) were not statistically different from that of control, meaning that BPA effect was reversible.
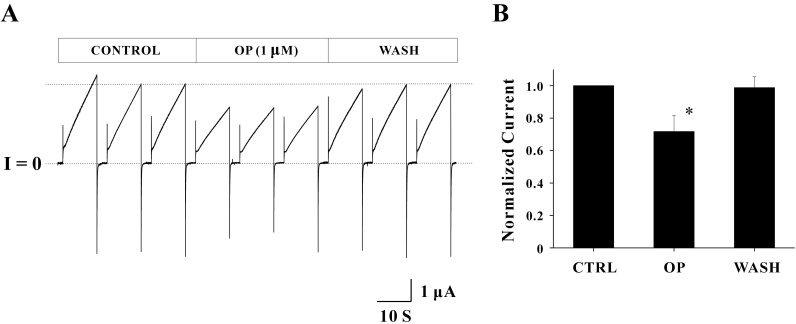
Fig. 3 The effect of OP (1 µM) on Cx46 hemichannel currents. (A) One set of records obtained from a single oocyte expressing Cx46 hemichannels are shown. The pulse protocol included VH=-70 mV, Vstep=+30 mV for 10 sec, and a total duration of 40 sec was used to each sweep. The average peak current in the control phase (1.60±0.13 µA) was reduced to 1.06±0.02 µA by applying 1 µM OP (middle three sweeps). The reduction rate of currents for this particular record was 33.72% of control. The inhibitory effect of OP was reversible by recovering reduced current back to 1.52±0.06 uA. Overall shapes for outward and inward currents were unchanged by OP. Current traces for about 25 seconds before the next sweep were omitted for better illustration. (B) All data obtained from the four independent experiments were analyzed by normalizing peak currents of each drug (or wash) phase to those of the control phase. Normalized peak currents inhibited by OP were 0.72±0.10 (n=4) and this current reduction (~28% of control) was statistically significant (*p<0.05). Inhibitory effect of OP was also reversible (0.99±0.07, n=4).
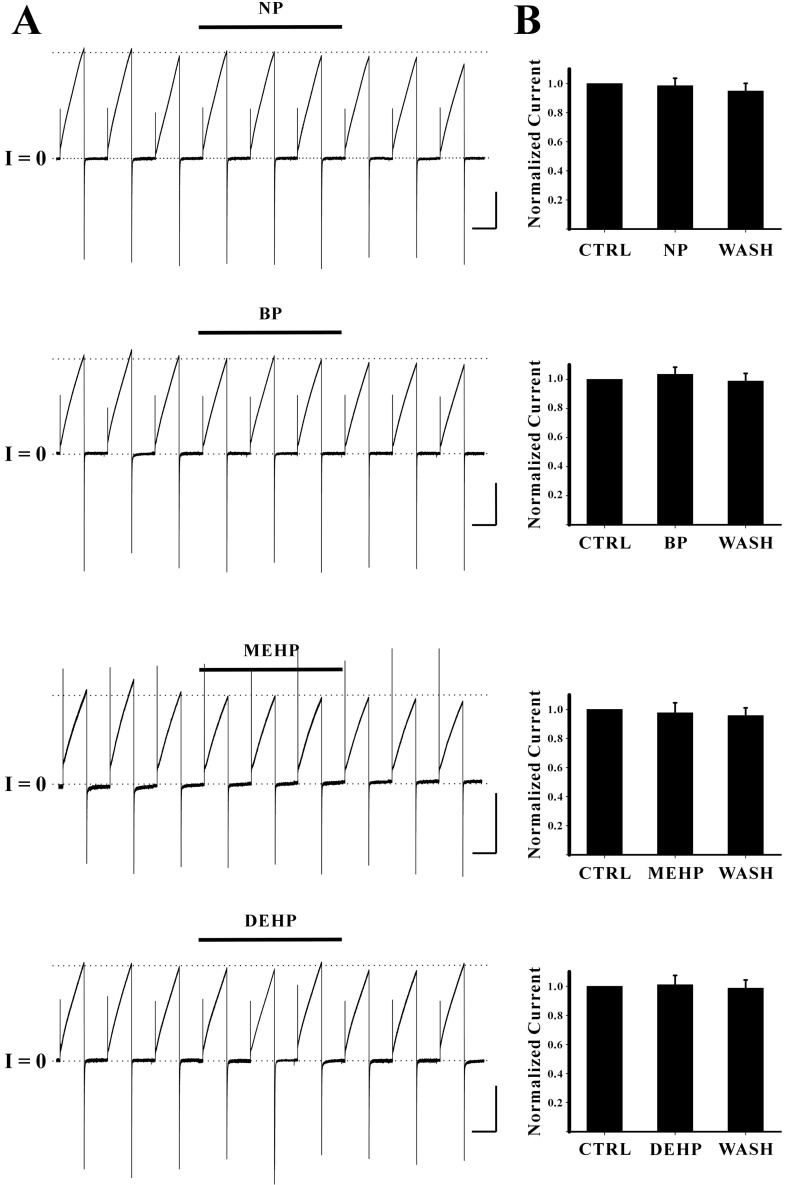
Fig. 4 The effects of other drugs on Cx46 hemichannel currents. (A) Representative current traces obtained from oocytes expressing Cx46 hemichannels with drug treatments are shown. The pulse protocol (VH=-70 mV, Vstep=+30 mV for 10 sec, and a total duration of 40 sec) was used for each sweep. The horizontal thick bars on the top of current traces indicate drug phases (from the top panel, NP, BP, MEHP, and DEHP, respectively). The average peak currents in control phase (initial three current traces of each record) were not changed by drug treatments indicated by dotted lines. Current traces and overall shapes for outward and inward currents of Cx46 hemichannels were not modified by these drugs. None of them appeared to block Cx46 hemichannel currents. Current traces for about 20 seconds before the next sweep were omitted for better illustration. Scale bars indicate 10 sec and 1 µA. (B) Normalized peak currents obtained from the independent experiments are shown. Normalized peak current of each phase was determined by the ratio of average peak currents of the drug (or wash) phase to those of the control phase. Normalized peak currents with drug treatments (0.99±0.05 for NP (n=3), 1.03±0.05 for BP (n=3), 0.98±0.07 for MEHP (n=4), 1.01±0.06 for DEHP (n=3), respectively) were not significantly different from those of control.
Reference
-
1. Harris AL. Emerging issues of connexin channels: biophysics fills the gap. Q Rev Biophys. 2001; 34:325–472. PMID: 11838236.2. Paul DL. Molecular cloning of cDNA for rat liver gap junction protein. J Cell Biol. 1986; 103:123–134. PMID: 3013898.
Article3. Söhl G, Willecke K. Gap junctions and the connexin protein family. Cardiovasc Res. 2004; 62:228–232. PMID: 15094343.4. Beyer EC, Paul DL, Goodenough DA. Connexin43: a protein from rat heart homologous to a gap junction protein from liver. J Cell Biol. 1987; 105:2621–2629. PMID: 2826492.
Article5. Kumar NM, Friend DS, Gilula NB. Synthesis and assembly of human beta 1 gap junctions in BHK cells by DNA transfection with the human beta 1 cDNA. J Cell Sci. 1995; 108:3725–3734. PMID: 8719879.
Article6. Hennemann H, Dahl E, White JB, Schwarz HJ, Lalley PA, Chang S, Nicholson BJ, Willecke K. Two gap junction genes, connexin 31.1 and 30.3, are closely linked on mouse chromosome 4 and preferentially expressed in skin. J Biol Chem. 1992; 267:17225–17233. PMID: 1512260.
Article7. Chang M, Werner R, Dahl G. A role for an inhibitory connexin in testis? Dev Biol. 1996; 175:50–56. PMID: 8608868.
Article8. Verselis VK, Ginter CS, Bargiello TA. Opposite voltage gating polarities of two closely related connexins. Nature. 1994; 368:348–351. PMID: 8127371.9. Oh S, Ri Y, Bennett MV, Trexler EB, Verselis VK, Bargiello TA. Changes in permeability caused by connexin 32 mutations underlie X-linked Charcot-Marie-Tooth disease. Neuron. 1997; 19:927–938. PMID: 9354338.
Article10. Paul DL, Ebihara L, Takemoto LJ, Swenson KI, Goodenough DA. Connexin46, a novel lens gap junction protein, induces voltage-gated currents in nonjunctional plasma membrane of Xenopus oocytes. J Cell Biol. 1991; 115:1077–1089. PMID: 1659572.
Article11. Trexler EB, Bennett MV, Bargiello TA, Verselis VK. Voltage gating and permeation in a gap junction hemichannel. Proc Natl Acad Sci U S A. 1996; 93:5836–5841. PMID: 8650179.
Article12. Diamanti-Kandarakis E, Bourguignon JP, Giudice LC, Hauser R, Prins GS, Soto AM, Zoeller RT, Gore AC. Endocrine-disrupting chemicals: an Endocrine Society scientific statement. Endocr Rev. 2009; 30:293–342. PMID: 19502515.
Article13. Wright CS, Becker DL, Lin JS, Warner AE, Hardy K. Stage-specific and differential expression of gap junctions in the mouse ovary: connexin-specific roles in follicular regulation. Reproduction. 2001; 121:77–88. PMID: 11226030.
Article14. Grazul-Bilska AT, Reynolds LP, Redmer DA. Gap junctions in the ovaries. Biol Reprod. 1997; 57:947–957. PMID: 9369157.15. Risley MS. Connexin gene expression in seminiferous tubules of the Sprague-Dawley rat. Biol Reprod. 2000; 62:748–754. PMID: 10684819.16. Vandenberg LN, Colborn T, Hayes TB, Heindel JJ, Jacobs DR Jr, Lee DH, Shioda T, Soto AM, vom Saal FS, Welshons WV, Zoeller RT, Myers JP. Hormones and endocrine-disrupting chemicals: low-dose effects and nonmonotonic dose responses. Endocr Rev. 2012; 33:378–455. PMID: 22419778.
Article17. Rottgen TS, Fancher IS, Asano S, Widlanski TS, Dick GM. Bisphenol A activates BK channels through effects on α and β1 subunits. Channels (Austin). 2014; 8:249–257.
Article18. O'Reilly AO, Eberhardt E, Weidner C, Alzheimer C, Wallace BA, Lampert A. Bisphenol A binds to the local anesthetic receptor site to block the human cardiac sodium channel. PLoS One. 2012; 7:e41667. PMID: 22848561.19. Deutschmann A, Hans M, Meyer R, Häberlein H, Swandulla D. Bisphenol A inhibits voltage-activated Ca2+ channels in vitro: mechanisms and structural requirements. Mol Pharmacol. 2013; 83:501–511. PMID: 23197648.20. Gao Q, Zhu T, Guo F, Huang S, Hu H, Feng R, Hao L. Nonylphenol, an environmental estrogen, affects voltage-gated K+ currents and L-type Ca2+ currents in a non-monotonic manner in GH3 pituitary cells. Toxicol Lett. 2013; 218:137–143. PMID: 23376477.21. Spray DC, Rozental R, Srinivas M. Prospects for rational development of pharmacological gap junction channel blockers. Curr Drug Targets. 2002; 3:455–464. PMID: 12448697.22. Boger DL, Patterson JE, Guan X, Cravatt BF, Lerner RA, Gilula NB. Chemical requirements for inhibition of gap junction communication by the biologically active lipid oleamide. Proc Natl Acad Sci U S A. 1998; 95:4810–4815. PMID: 9560184.
Article23. Srinivas M, Hopperstad MG, Spray DC. Quinine blocks specific gap junction channel subtypes. Proc Natl Acad Sci U S A. 2001; 98:10942–10947. PMID: 11535816.
Article24. Kim DY, Jung CS. Gap junction contributions to the goldfish electroretinogram at the photopic illumination level. Korean J Physiol Pharmacol. 2012; 16:219–224. PMID: 22802705.
Article25. Oh S, Abrams CK, Verselis VK, Bargiello TA. Stoichiometry of transjunctional voltage-gating polarity reversal by a negative charge substitution in the amino terminus of a connexin32 chimera. J Gen Physiol. 2000; 116:13–31. PMID: 10871637.
Article26. Rubin JB, Verselis VK, Bennett MV, Bargiello TA. Molecular analysis of voltage dependence of heterotypic gap junctions formed by connexins 26 and 32. Biophys J. 1992; 62:183–193. PMID: 1376166.
Article27. Trexler EB, Bukauskas FF, Kronengold J, Bargiello TA, Verselis VK. The first extracellular loop domain is a major determinant of charge selectivity in connexin46 channels. Biophys J. 2000; 79:3036–3051. PMID: 11106610.
Article28. Derouiche S, Warnier M, Mariot P, Gosset P, Mauroy B, Bonnal JL, Slomianny C, Delcourt P, Prevarskaya N, Roudbaraki M. Bisphenol A stimulates human prostate cancer cell migration via remodelling of calcium signalling. Springerplus. 2013; 2:54. PMID: 23450760.
Article29. Chung JY, Kim JY, Kim YJ, Jung SJ, Park JE, Lee SG, Kim JT, Oh S, Lee CJ, Yoon YD, Yoo YH, Kim JM. Cellular defense mechanisms against benzo[a]pyrene in testicular Leydig cells: implications of p53, aryl-hydrocarbon receptor, and cytochrome P450 1A1 status. Endocrinology. 2007; 148:6134–6144. PMID: 17884947.
Article30. Kotula-Balak M, Chojnacka K, Hejmej A, Galas J, Satola M, Bilinska B. Does 4-tert-octylphenol affect estrogen signaling pathways in bank vole Leydig cells and tumor mouse Leydig cells in vitro? Reprod Toxicol. 2013; 39:6–16. PMID: 23557686.
Article31. Liu PS, Liu GH, Chao WL. Effects of nonylphenol on the calcium signal and catecholamine secretion coupled with nicotinic acetylcholine receptors in bovine adrenal chromaffin cells. Toxicology. 2008; 244:77–85. PMID: 18093714.
Article32. Ambruosi B, Uranio MF, Sardanelli AM, Pocar P, Martino NA, Paternoster MS, Amati F, Dell'Aquila ME. In vitro acute exposure to DEHP affects oocyte meiotic maturation, energy and oxidative stress parameters in a large animal model. PLoS One. 2011; 6:e27452. PMID: 22076161.
Article33. Chauvigné F, Plummer S, Lesné L, Cravedi JP, Dejucq-Rainsford N, Fostier A, Jégou B. Mono-(2-ethylhexyl) phthalate directly alters the expression of Leydig cell genes and CYP17 lyase activity in cultured rat fetal testis. PLoS One. 2011; 6:e27172. PMID: 22087261.
Article34. Kado RT, Baud C. The rise and fall of electrical excitability in the oocyte of Xenopus laevis. J Physiol (Paris). 1981; 77:1113–1117. PMID: 6286961.35. Gil Z, Magleby KL, Silberberg SD. Membrane-pipette interactions underlie delayed voltage activation of mechanosensitive channels in Xenopus oocytes. Biophys J. 1999; 76:3118–3127. PMID: 10354436.
Article36. Rettinger J. Novel properties of the depolarization-induced endogenous sodium conductance in the Xenopus laevis oocyte. Pflugers Arch. 1999; 437:917–924. PMID: 10370071.
Article37. Trexler EB, Bukauskas FF, Bennett MV, Bargiello TA, Verselis VK. Rapid and direct effects of pH on connexins revealed by the connexin46 hemichannel preparation. J Gen Physiol. 1999; 113:721–742. PMID: 10228184.38. Hamill OP, McBride DW Jr. Rapid adaptation of single mechanosensitive channels in Xenopus oocytes. Proc Natl Acad Sci U S A. 1992; 89:7462–7466. PMID: 1380158.
Article39. Unger VM, Kumar NM, Gilula NB, Yeager M. Three-dimensional structure of a recombinant gap junction membrane channel. Science. 1999; 283:1176–1180. PMID: 10024245.
Article40. Maeda S, Nakagawa S, Suga M, Yamashita E, Oshima A, Fujiyoshi Y, Tsukihara T. Structure of the connexin 26 gap junction channel at 3.5 A resolution. Nature. 2009; 458:597–602. PMID: 19340074.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Adverse effects of 4-tert-octylphenol on the production of oxytocin and hCG in pregnant rats
- Mechanisms of TERT Reactivation and Its Interaction with BRAFV600E
- Male preference for TERT alterations and HBV integration in young-age HBV-related HCC: implications for sex disparity
- Analysis of the Cancer Genome Atlas Data to Determine the Prognostic Value of GABPB1L and TERT in Glioblastoma
- A Survey of Disinfection Methods for Formula Bottle
