Protective Effect of Delta-Like 1 Homolog Against Muscular Atrophy in a Mouse Model
- Affiliations
-
- 1Department of Molecular, Cellular and Cancer Biology, Graduate School of Medical Science, Brain Korea 21 Project, Yonsei University College of Medicine, Seoul, Korea
- 2Department of Internal Medicine, Yonsei University College of Medicine, Seoul, Korea
- 3Institute of Endocrine Research, Yonsei University College of Medicine, Seoul, Korea
- 4Y-Biologics, Inc., Daejeon, Korea
- KMID: 2532868
- DOI: http://doi.org/10.3803/EnM.2022.1446
Abstract
- Background
Muscle atrophy is caused by an imbalance between muscle growth and wasting. Delta-like 1 homolog (DLK1), a protein that modulates adipogenesis and muscle development, is a crucial regulator of myogenic programming. Thus, we investigated the effect of exogenous DLK1 on muscular atrophy.
Methods
We used muscular atrophy mouse model induced by dexamethasone (Dex). The mice were randomly divided into three groups: (1) control group, (2) Dex-induced muscle atrophy group, and (3) Dex-induced muscle atrophy group treated with DLK1. The effects of DLK1 were also investigated in an in vitro model using C2C12 myotubes.
Results
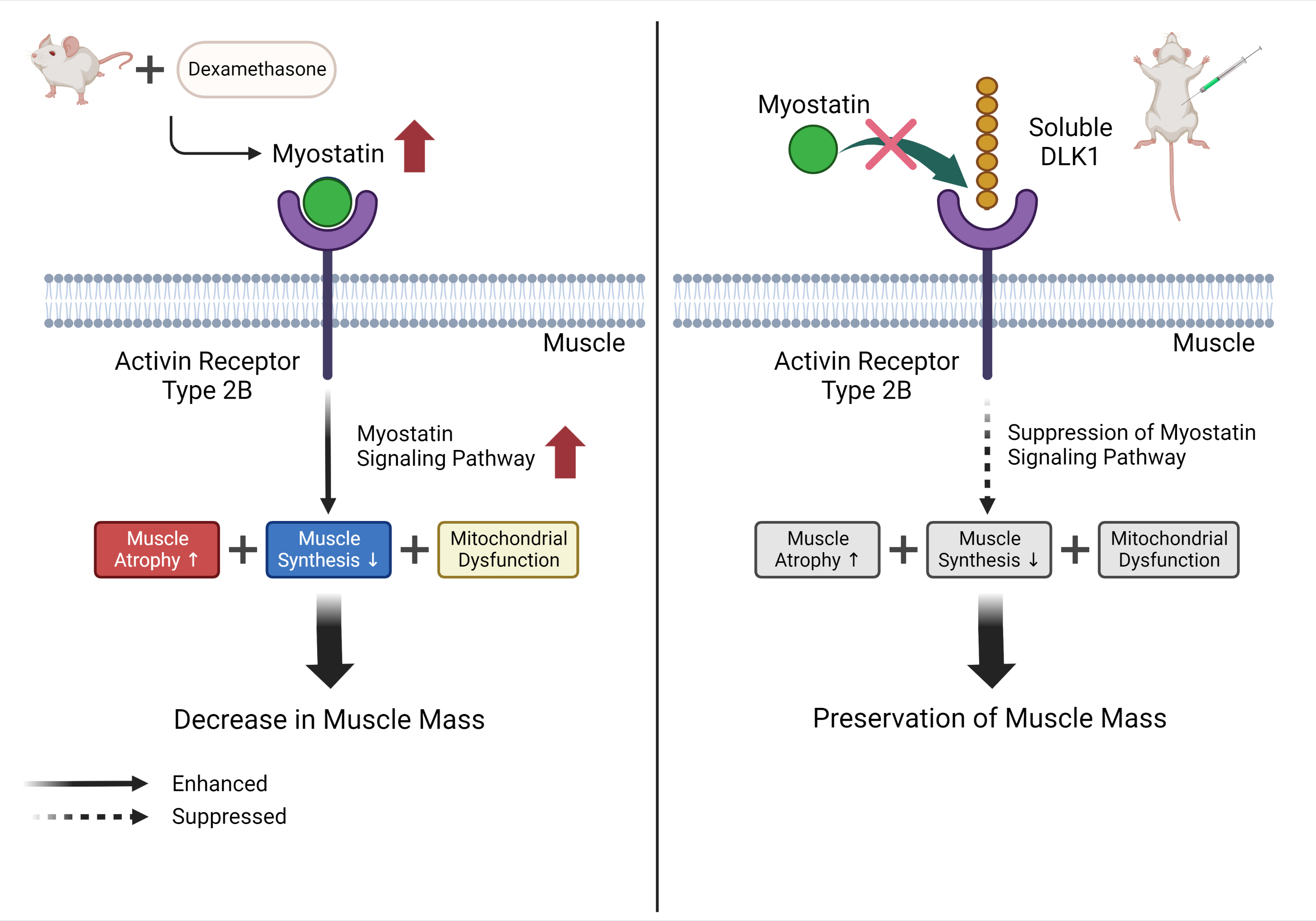
Dex-induced muscular atrophy in mice was associated with increased expression of muscle atrophy markers and decreased expression of muscle differentiation markers, while DLK1 treatment attenuated these degenerative changes together with reduced expression of the muscle growth inhibitor, myostatin. In addition, electron microscopy revealed that DLK1 treatment improved mitochondrial dynamics in the Dex-induced atrophy model. In the in vitro model of muscle atrophy, normalized expression of muscle differentiation markers by DLK1 treatment was mitigated by myostatin knockdown, implying that DLK1 attenuates muscle atrophy through the myostatin pathway.
Conclusion
DLK1 treatment inhibited muscular atrophy by suppressing myostatin-driven signaling and improving mitochondrial biogenesis. Thus, DLK1 might be a promising candidate to treat sarcopenia, characterized by muscle atrophy and degeneration.
Keyword
Figure
-
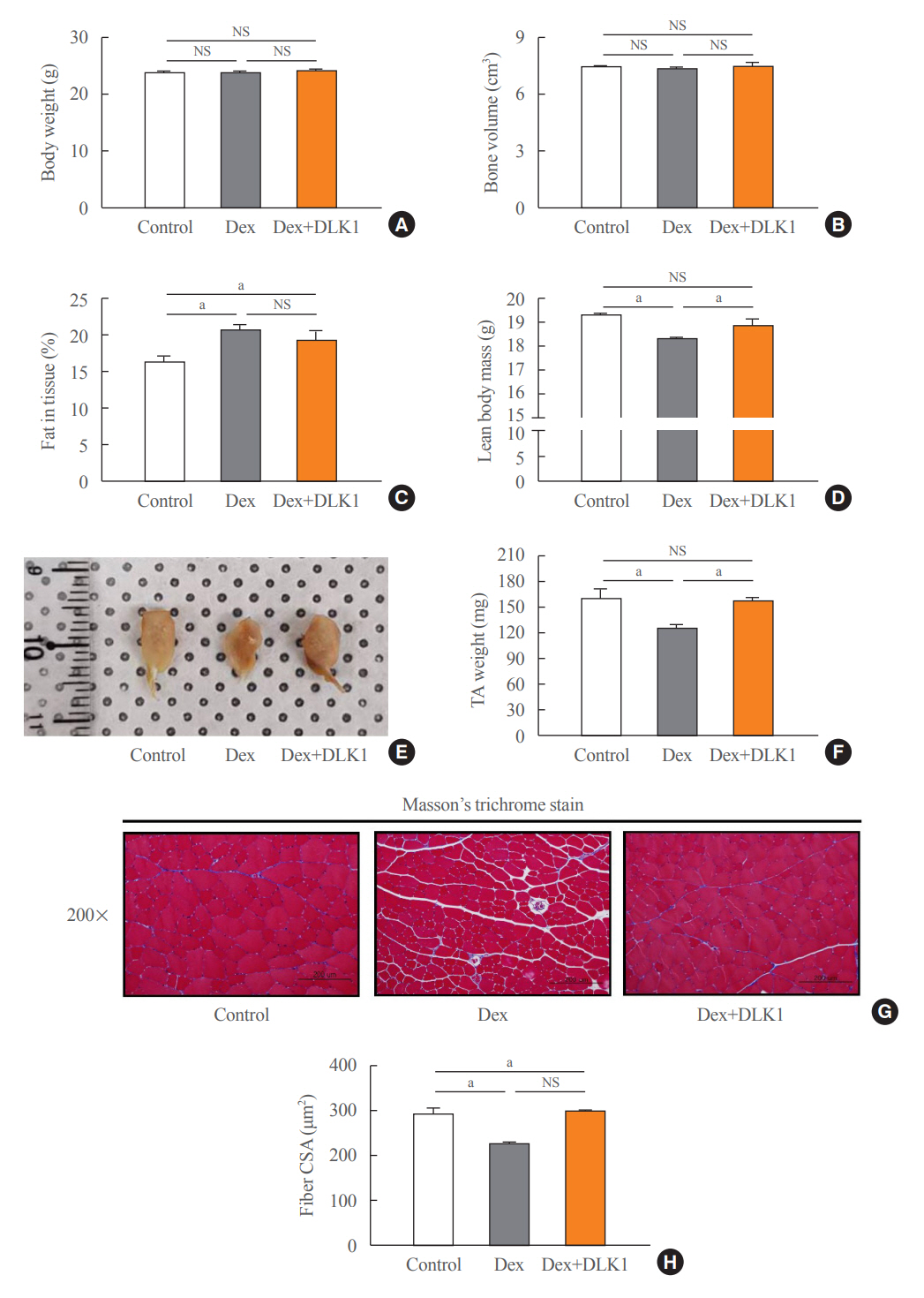
Fig. 1. Effects of delta-like 1 homolog (DLK1) on body composition and histopathological morphology of tibialis anterior muscle in the dexamethasone-induced muscle atrophy mouse model. (A, B, C, D) Effects of DLK1 on body composition in dexamethasone-induced tibialis anterior (TA) skeletal muscle atrophy mice using dual energy X-ray absorptiometry (n=4 per each group). (E) Size comparison of TA muscles from a mouse hindlimb. (F) TA muscle weights (n=6 per each group). (G) Representative histopathological images of TA muscle (200× magnification; scale bar 200 μm) using Masson’s trichrome stain. (H) Mean cross-sectional area of the TA muscle (μm2). Results are presented as mean±standard error of the mean. Treatment groups were as follows: control=oral distilled water and intraperitoneal (IP) phosphate buffered saline (PBS); dexamethasone (Dex)=oral Dex (1 mg/kg) and IP PBS; Dex+DLK1=oral Dex (1 mg/kg) and IP DLK1 (0.8 mg/kg/day). CSA, cross-sectional area; NS, non-significant. aP<0.05 was considered statistically significant.
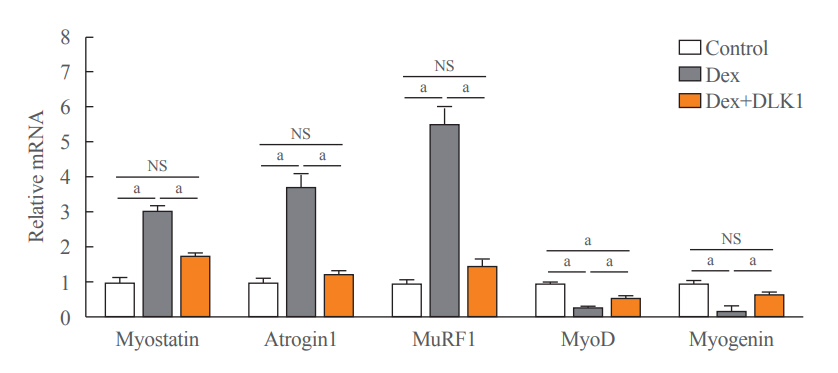
Fig. 2. Effects of delta-like 1 homolog (DLK1) on muscle atrophic and myogenic factors in the dexamethasone-induced muscle atrophy mouse model. A graph showing relative mRNA expression of muscle atrophic (myostatin, atrogin1, and muscle-specific RING finger protein 1 [MuRF1]) and myogenic (myogenic differentiation [MyoD] and myogenin) factors in the mouse tibialis anterior (TA) muscle. Results are presented as mean±standard error of the mean. Treatment groups were as follows: control=oral distilled water and intraperitoneal (IP) phosphate buffered saline (PBS); dexamethasone (Dex)=oral Dex (1 mg/kg) and IP PBS; Dex+DLK1=oral Dex (1 mg/kg) and IP DLK1 (0.8 mg/kg/day). NS, non-significant. aP<0.05 was considered statistically significant.
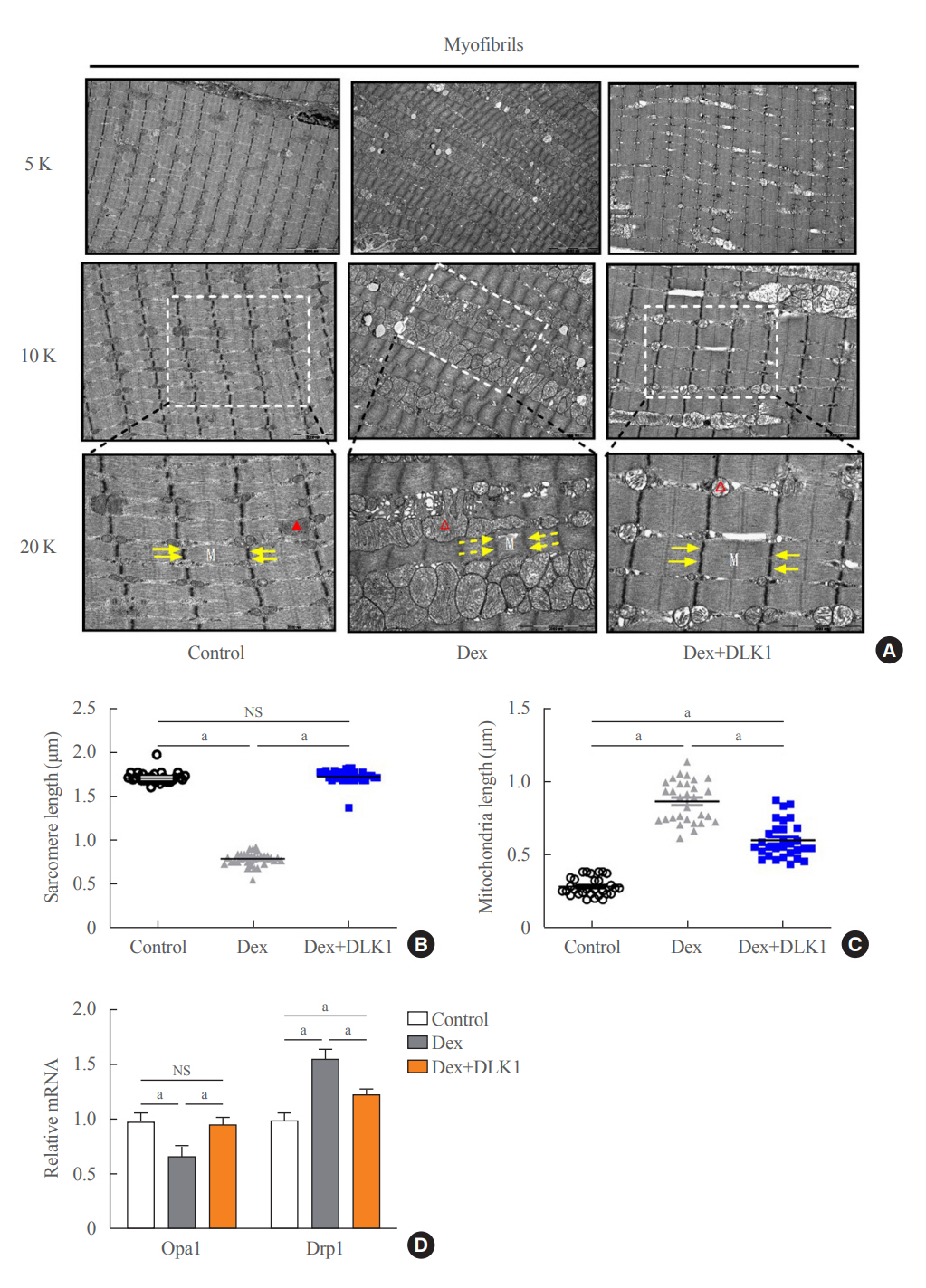
Fig. 3. Effects of delta-like 1 homolog (DLK1) on tibialis anterior muscle morphology and mitochondrial biogenesis markers in the dexamethasone-induced muscle atrophy mouse model. (A) Representative electron microscopic images of myofibrils within the tibialis anterior (TA) muscle (5,000×, 10,000×, and 20,000× magnification; M=myofilament, yellow arrow=Z line, red arrow=mitochondria, dashed arrow=a broken structure, hollow red arrow=a swollen structure). Graphs showing lengths of (B) sarcomeres and (C) mitochondria in TA muscle cells (n=30 per each group). (D) Relative mRNA expression of mitochondrial fusion (Opa1) and fission (Drp1) markers (n=10 per each group). Results are presented as mean±standard error of the mean. Treatment groups were as follows: control=oral distilled water and intraperitoneal (IP) phosphate buffered saline (PBS); dexamethasone (Dex)=oral Dex (1 mg/kg) and IP PBS; Dex+DLK1=oral Dex (1 mg/kg) and IP DLK1 (0.8 mg/kg/day). NS, non-significant. aP<0.05 was considered statistically significant.
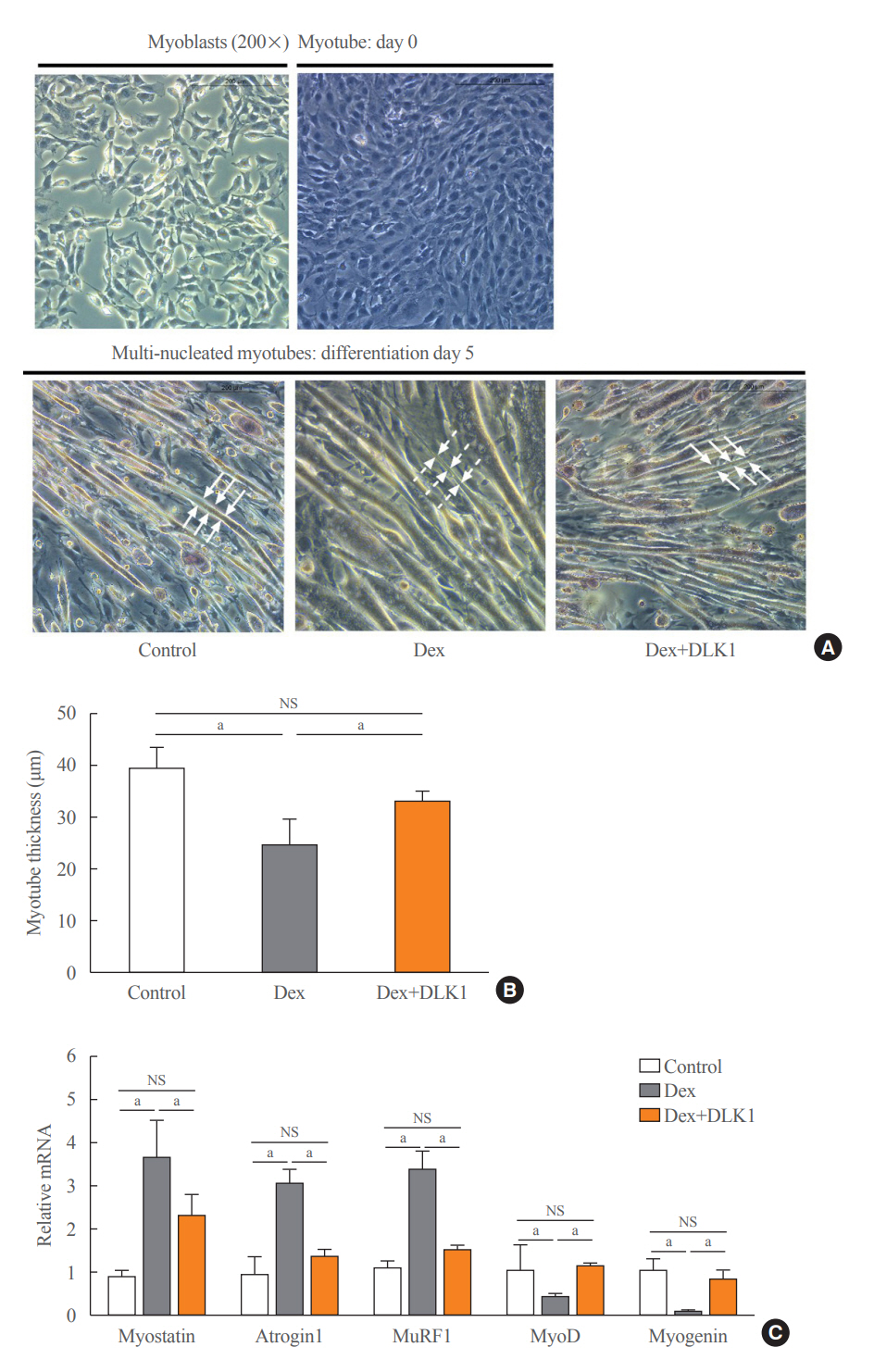
Fig. 4. Effect of delta-like 1 homolog (DLK1) on dexamethasone-induced myotube atrophy. (A) Representative histopathological images of H&E staining showing myoblast (Day 0) and differentiated multi-nucleated myotubes (Day 5) according to the type of differentiation medium. (B) Thickness of myotubes (µm). (C) Relative mRNA expression of muscle atrophic (myostatin, atrogin1, and muscle-specific RING finger protein 1 [MuRF1]) and myogenic (myogenic differentiation [MyoD] and myogenin) factors in myotubes. Results are presented as mean±standard error of the mean. Myotube cultures were divided into three groups on differentiation Day 4 and cultured in different media for 24 hours: control=Dulbecco’s Modified Eagle Medium (DMEM)+ethanol (0.02% v/v); dexamethasone (Dex)=DMEM+Dex (10 μM dissolved in ethanol); and Dex+DLK1=DMEM+Dex (10 μM dissolved in ethanol)+DLK1 (5.0 μg/mL). NS, non-significant. aP<0.05 was considered statistically significant.
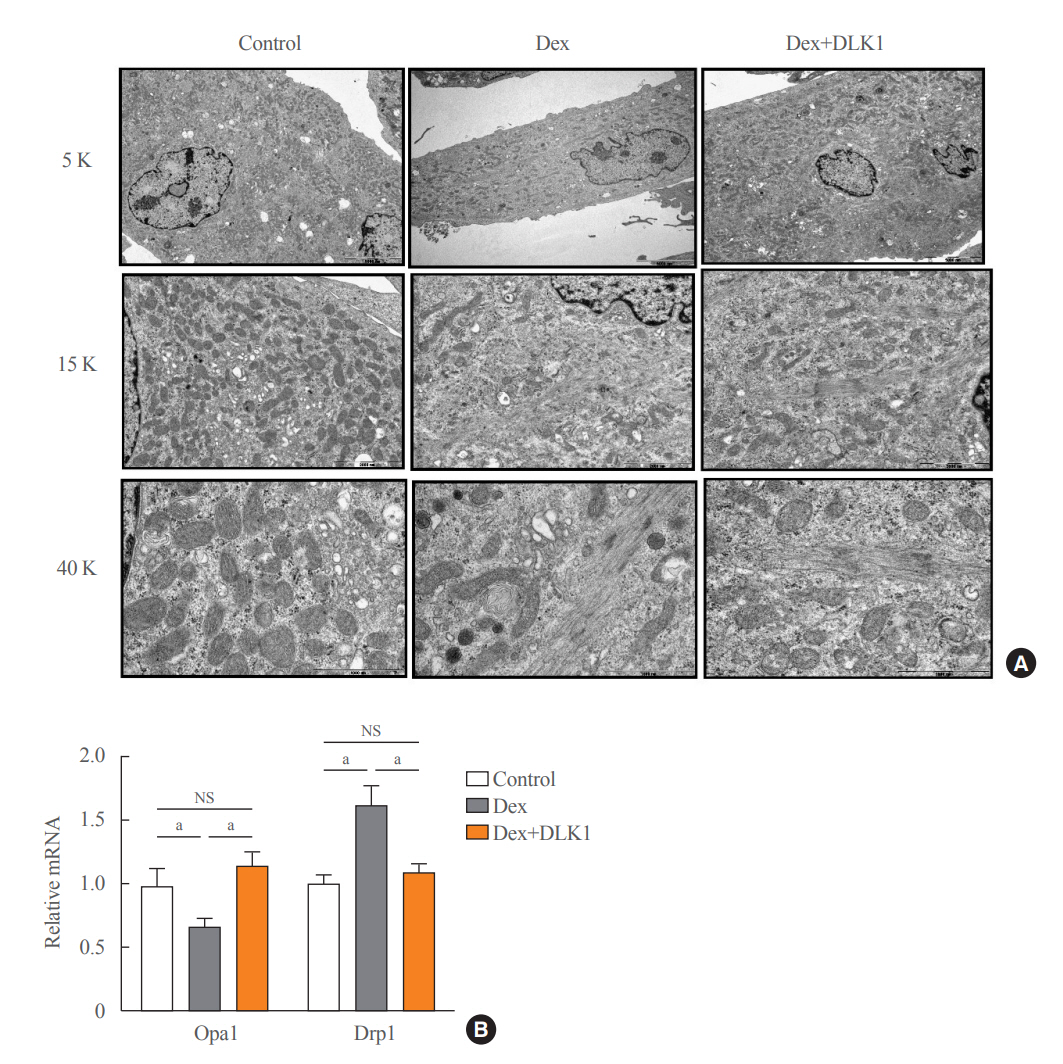
Fig. 5. Effects of delta-like 1 homolog (DLK1) on mitochondrial biogenesis markers in dexamethasone-induced myotube atrophy. (A) Representative electron microscopic images of myotube cells (5,000×, 15,000×, and 40,000× magnification). (B) Relative mRNA expression of mitochondrial fusion (Opa1) and fission (Drp1) markers (n=10 per each group). Results are presented as mean±standard error of the mean. Myotube cultures were divided into three groups on differentiation Day 4 and cultured in different media for 24 hours: control=Dulbecco’s Modified Eagle Medium (DMEM)+ethanol (0.02% v/v); dexamethasone (Dex)=DMEM+Dex (10 μM dissolved in ethanol); and Dex+DLK1=DMEM+Dex (10 μM dissolved in ethanol)+DLK1 (5.0 μg/mL). NS, non-significant. aP<0.05 was considered statistically significant.
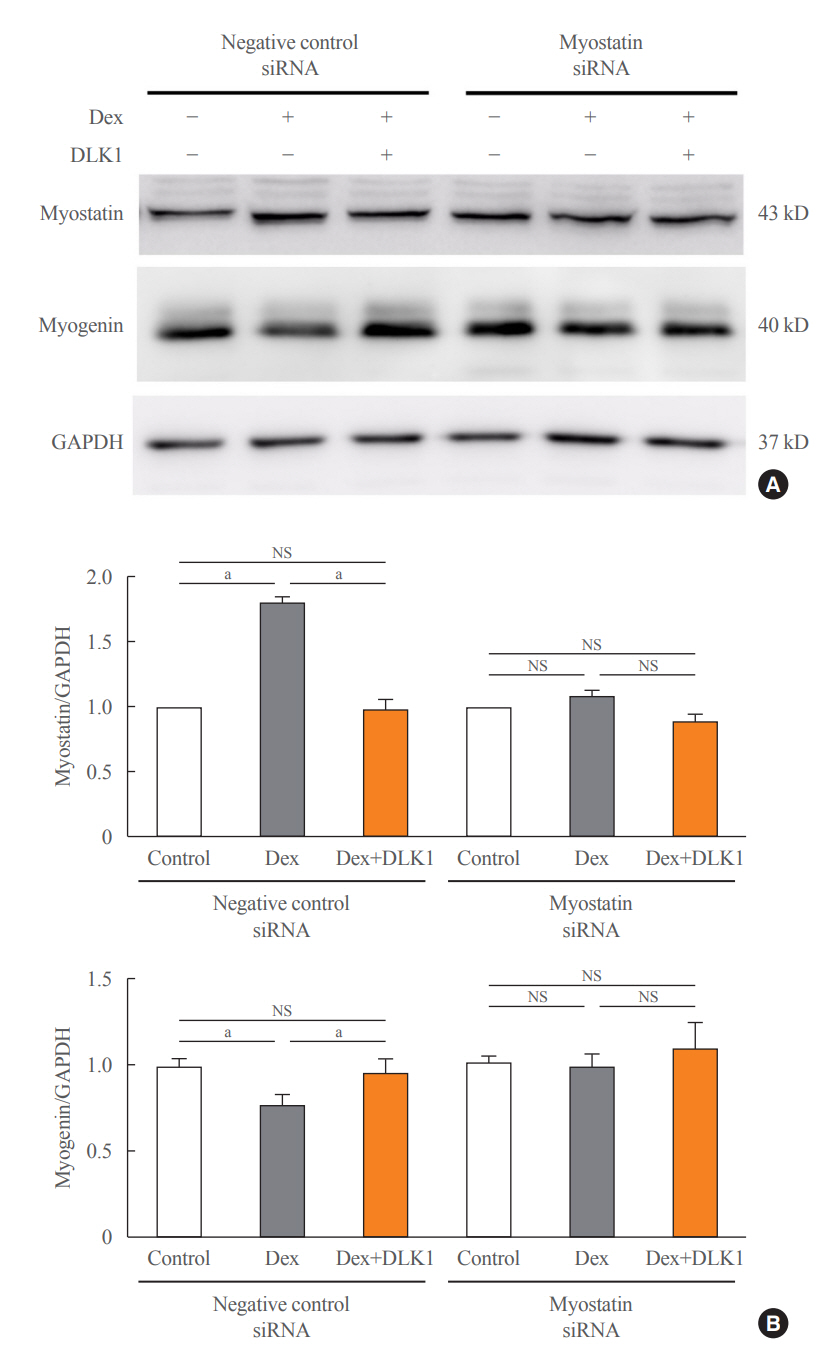
Fig. 6. Effect of delta-like 1 homolog (DLK1) on muscle-related markers in atrophied myotubes with or without knockdown of myostatin expression. (A) Western blot images with (B) quantification of myostatin and myogenin protein after transfection with small interfering ribonucleic acids (siRNAs) of negative control or myostatin (n=3 to 6 per group). Results are presented as mean±standard error of the mean. Transfected myotube cultures were divided into three groups on differentiation Day 4 and cultured in different media for 24 hours: control=Dulbecco’s Modified Eagle Medium (DMEM)+ethanol (0.02% v/v); dexamethasone (Dex)=DMEM+Dex (10 μM dissolved in ethanol); and Dex+DLK1=DMEM+Dex (10 μM dissolved in ethanol)+DLK1 (5.0 μg/mL). GAPDH, glyceraldehyde-3-phosphate dehydrogenase; NS, non-significant. aP<0.05 was considered statistically significant.
Reference
-
1. Fanzani A, Conraads VM, Penna F, Martinet W. Molecular and cellular mechanisms of skeletal muscle atrophy: an update. J Cachexia Sarcopenia Muscle. 2012; 3:163–79.
Article2. Saini A, Faulkner S, Al-Shanti N, Stewart C. Powerful signals for weak muscles. Ageing Res Rev. 2009; 8:251–67.
Article3. Waddell JN, Zhang P, Wen Y, Gupta SK, Yevtodiyenko A, Schmidt JV, et al. Dlk1 is necessary for proper skeletal muscle development and regeneration. PLoS One. 2010; 5:e15055.
Article4. Lee YH, Yun MR, Kim HM, Jeon BH, Park BC, Lee BW, et al. Exogenous administration of DLK1 ameliorates hepatic steatosis and regulates gluconeogenesis via activation of AMPK. Int J Obes (Lond). 2016; 40:356–65.
Article5. Morrison-Nozik A, Anand P, Zhu H, Duan Q, Sabeh M, Prosdocimo DA, et al. Glucocorticoids enhance muscle endurance and ameliorate Duchenne muscular dystrophy through a defined metabolic program. Proc Natl Acad Sci U S A. 2015; 112:E6780–9.
Article6. Ma K, Mallidis C, Bhasin S, Mahabadi V, Artaza J, Gonzalez-Cadavid N, et al. Glucocorticoid-induced skeletal muscle atrophy is associated with upregulation of myostatin gene expression. Am J Physiol Endocrinol Metab. 2003; 285:E363–71.7. Menconi M, Gonnella P, Petkova V, Lecker S, Hasselgren PO. Dexamethasone and corticosterone induce similar, but not identical, muscle wasting responses in cultured L6 and C2C12 myotubes. J Cell Biochem. 2008; 105:353–64.
Article8. Wang R, Jiao H, Zhao J, Wang X, Lin H. Glucocorticoids enhance muscle proteolysis through a myostatin-dependent pathway at the early stage. PLoS One. 2016; 11:e0156225.
Article9. Langone F, Cannata S, Fuoco C, Lettieri Barbato D, Testa S, Nardozza AP, et al. Metformin protects skeletal muscle from cardiotoxin induced degeneration. PLoS One. 2014; 9:e114018.
Article10. Costamagna D, Costelli P, Sampaolesi M, Penna F. Role of Inflammation in muscle homeostasis and myogenesis. Mediators Inflamm. 2015; 2015:805172.
Article11. Rodriguez J, Vernus B, Chelh I, Cassar-Malek I, Gabillard JC, Hadj Sassi A, et al. Myostatin and the skeletal muscle atrophy and hypertrophy signaling pathways. Cell Mol Life Sci. 2014; 71:4361–71.
Article12. Rebbapragada A, Benchabane H, Wrana JL, Celeste AJ, Attisano L. Myostatin signals through a transforming growth factor beta-like signaling pathway to block adipogenesis. Mol Cell Biol. 2003; 23:7230–42.
Article13. Morissette MR, Stricker JC, Rosenberg MA, Buranasombati C, Levitan EB, Mittleman MA, et al. Effects of myostatin deletion in aging mice. Aging Cell. 2009; 8:573–83.
Article14. Chen JL, Walton KL, Hagg A, Colgan TD, Johnson K, Qian H, et al. Specific targeting of TGF-β family ligands demonstrates distinct roles in the regulation of muscle mass in health and disease. Proc Natl Acad Sci U S A. 2017; 114:E5266–75.
Article15. Han HQ, Zhou X, Mitch WE, Goldberg AL. Myostatin/activin pathway antagonism: molecular basis and therapeutic potential. Int J Biochem Cell Biol. 2013; 45:2333–47.
Article16. Liu M, Hammers DW, Barton ER, Sweeney HL. Activin receptor type IIB inhibition improves muscle phenotype and function in a mouse model of spinal muscular atrophy. PLoS One. 2016; 11:e0166803.
Article17. Lach-Trifilieff E, Minetti GC, Sheppard K, Ibebunjo C, Feige JN, Hartmann S, et al. An antibody blocking activin type II receptors induces strong skeletal muscle hypertrophy and protects from atrophy. Mol Cell Biol. 2014; 34:606–18.
Article18. Langley B, Thomas M, Bishop A, Sharma M, Gilmour S, Kambadur R. Myostatin inhibits myoblast differentiation by down-regulating MyoD expression. J Biol Chem. 2002; 277:49831–40.
Article19. LeBrasseur NK, Schelhorn TM, Bernardo BL, Cosgrove PG, Loria PM, Brown TA. Myostatin inhibition enhances the effects of exercise on performance and metabolic outcomes in aged mice. J Gerontol A Biol Sci Med Sci. 2009; 64:940–8.
Article20. Gill JF, Delezie J, Santos G, McGuirk S, Schnyder S, Frank S, et al. Peroxisome proliferator-activated receptor γ coactivator 1α regulates mitochondrial calcium homeostasis, sarcoplasmic reticulum stress, and cell death to mitigate skeletal muscle aging. Aging Cell. 2019; 18:e12993.
Article21. Baldelli S, Aquilano K, Ciriolo MR. PGC-1α buffers ROS-mediated removal of mitochondria during myogenesis. Cell Death Dis. 2014; 5:e1515.
Article22. Schakman O, Gilson H, Thissen JP. Mechanisms of glucocorticoid-induced myopathy. J Endocrinol. 2008; 197:1–10.
Article23. Bodine SC, Latres E, Baumhueter S, Lai VK, Nunez L, Clarke BA, et al. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science. 2001; 294:1704–8.
Article24. Hernandez-Hernandez JM, Garcia-Gonzalez EG, Brun CE, Rudnicki MA. The myogenic regulatory factors, determinants of muscle development, cell identity and regeneration. Semin Cell Dev Biol. 2017; 72:10–8.
Article25. Shintaku J, Peterson JM, Talbert EE, Gu JM, Ladner KJ, Williams DR, et al. MyoD regulates skeletal muscle oxidative metabolism cooperatively with alternative NF-κB. Cell Rep. 2016; 17:514–26.
Article26. Flynn JM, Meadows E, Fiorotto M, Klein WH. Myogenin regulates exercise capacity and skeletal muscle metabolism in the adult mouse. PLoS One. 2010; 5:e13535.
Article27. Hasty P, Bradley A, Morris JH, Edmondson DG, Venuti JM, Olson EN, et al. Muscle deficiency and neonatal death in mice with a targeted mutation in the myogenin gene. Nature. 1993; 364:501–6.
Article28. Ma K, Mallidis C, Artaza J, Taylor W, Gonzalez-Cadavid N, Bhasin S. Characterization of 5’-regulatory region of human myostatin gene: regulation by dexamethasone in vitro. Am J Physiol Endocrinol Metab. 2001; 281:E1128–36.
Article29. Ronning SB. Myogenesis: methods and protocols. New York: Springer New York;2019. Chapter 13, A siRNA mediated screen during C2C12 myogenesis. 229–43.30. Elkina Y, von Haehling S, Anker SD, Springer J. The role of myostatin in muscle wasting: an overview. J Cachexia Sarcopenia Muscle. 2011; 2:143–51.
Article31. Forcina L, Cosentino M, Musaro A. Mechanisms regulating muscle regeneration: insights into the interrelated and time-dependent phases of tissue healing. Cells. 2020; 9:1297.
Article32. Yan Z, Choi S, Liu X, Zhang M, Schageman JJ, Lee SY, et al. Highly coordinated gene regulation in mouse skeletal muscle regeneration. J Biol Chem. 2003; 278:8826–36.
Article33. Bonaldo P, Sandri M. Cellular and molecular mechanisms of muscle atrophy. Dis Model Mech. 2013; 6:25–39.
Article34. Iqbal S, Hood DA. The role of mitochondrial fusion and fission in skeletal muscle function and dysfunction. Front Biosci (Landmark Ed). 2015; 20:157–72.
Article35. Tezze C, Romanello V, Desbats MA, Fadini GP, Albiero M, Favaro G, et al. Age-associated loss of OPA1 in muscle impacts muscle mass, metabolic homeostasis, systemic inflammation, and epithelial senescence. Cell Metab. 2017; 25:1374–89. e6.
Article36. Favaro G, Romanello V, Varanita T, Andrea Desbats M, Morbidoni V, Tezze C, et al. DRP1-mediated mitochondrial shape controls calcium homeostasis and muscle mass. Nat Commun. 2019; 10:2576.
Article37. Liu YJ, McIntyre RL, Janssens GE, Houtkooper RH. Mitochondrial fission and fusion: a dynamic role in aging and potential target for age-related disease. Mech Ageing Dev. 2020; 186:111212.
Article38. Del Campo A, Jaimovich E, Tevy MF. Mitochondria in the aging muscles of flies and mice: new perspectives for old characters. Oxid Med Cell Longev. 2016; 2016:9057593.39. Del Campo A, Contreras-Hernandez I, Castro-Sepulveda M, Campos CA, Figueroa R, Tevy MF, et al. Muscle function decline and mitochondria changes in middle age precede sarcopenia in mice. Aging (Albany NY). 2018; 10:34–55.
Article40. Sayed RK, de Leonardis EC, Guerrero-Martinez JA, Rahim I, Mokhtar DM, Saleh AM, et al. Identification of morphological markers of sarcopenia at early stage of aging in skeletal muscle of mice. Exp Gerontol. 2016; 83:22–30.
Article41. Walston JD. Sarcopenia in older adults. Curr Opin Rheumatol. 2012; 24:623–7.
Article42. Crupi AN, Nunnelee JS, Taylor DJ, Thomas A, Vit JP, Riera CE, et al. Oxidative muscles have better mitochondrial homeostasis than glycolytic muscles throughout life and maintain mitochondrial function during aging. Aging (Albany NY). 2018; 10:3327–52.
Article43. Schiaffino S, Dyar KA, Ciciliot S, Blaauw B, Sandri M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013; 280:4294–314.
Article44. Jensen CH, Kosmina R, Ryden M, Baun C, Hvidsten S, Andersen MS, et al. The imprinted gene delta like non-canonical notch ligand 1 (Dlk1) associates with obesity and triggers insulin resistance through inhibition of skeletal muscle glucose uptake. EBioMedicine. 2019; 46:368–80.
Article45. Wang M, Tan Y, Shi Y, Wang X, Liao Z, Wei P. Diabetes and sarcopenic obesity: pathogenesis, diagnosis, and treatments. Front Endocrinol (Lausanne). 2020; 11:568.
Article46. Bonetto A, Andersson DC, Waning DL. Assessment of muscle mass and strength in mice. Bonekey Rep. 2015; 4:732.
Article47. Mankhong S, Kim S, Moon S, Kwak HB, Park DH, Kang JH. Experimental models of sarcopenia: bridging molecular mechanism and therapeutic strategy. Cells. 2020; 9:1385.
Article48. Jang HC. Sarcopenia, frailty, and diabetes in older adults. Diabetes Metab J. 2016; 40:182–9.
Article49. Romanello V, Sandri M. Mitochondrial quality control and muscle mass maintenance. Front Physiol. 2016; 6:422.
Article50. Djafarzadeh S, Jakob SM. High-resolution respirometry to assess mitochondrial function in permeabilized and intact cells. J Vis Exp. 2017; 120:54985.
Article51. Dent JR, Hetrick B, Tahvilian S, Sathe A, Greyslak K, LaBarge SA, et al. Skeletal muscle mitochondrial function and exercise capacity are not impaired in mice with knockout of STAT3. J Appl Physiol (1985). 2019; 127:1117–27.
Article
