Impact of Nicotine Exposure on Hair Cell Toxicity and Embryotoxicity During Zebrafish Development
- Affiliations
-
- 1Department of Otorhinolaryngology-Head and Neck Surgery, Kyungpook National University School of Medicine, Daegu, Korea.
- 2Department of Otorhinolaryngology-Head and Neck Surgery, Korea University College of Medicine, Seoul, Korea. mednlaw@korea.ac.kr
- 3Biomedical Research Center, Korea University Ansan Hospital, Ansan, Korea.
- KMID: 2412668
- DOI: http://doi.org/10.21053/ceo.2017.00857
Abstract
OBJECTIVES
Nicotine has various adverse effects including negative impacts associated with maternal exposure. In the current study, we examined nicotine-induced damage of hair cells and embryotoxicity during zebrafish development.
METHODS
Zebrafish embryos were exposed to nicotine at several concentrations (5, 10, 20, and 40 μM) and embryotoxicity were evaluated at 72 hours, including hatching rate, mortality, teratogenicity rate, and heart rate. Hair cells within the supraorbital (SO1 and SO2), otic (O1), and occipital (OC1) neuromasts were identified at 120 hours. Apoptosis and mitochondrial damage of hair cells were analyzed using TUNEL (terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling) and DASPEI (2-[4-(dimethylamino)styryl]-N-ethylpyridinium iodide) assays, respectively, and changes of ultrastructure were observed by scanning electron microscopy.
RESULTS
The control group without nicotine appeared normal with overall mortality and teratogenicity rate < 5%. The hatching rate and mortality rate was not significantly different according to nicotine concentration (n=400 each). The abnormal morphology rate (n=400) increased and heart rate (n=150) decreased with increasing nicotine concentration (P < 0.05). Nicotine-induced hair cell damage significantly increased as nicotine concentration increased. A significantly greater number of TUNEL-positive cells (P < 0.01) and markedly smaller DASPEI area (P < 0.01) were shown as nicotine concentration increased.
CONCLUSION
The current results suggest that nicotine induces dose-dependent hair cell toxicity in embryos by promoting apoptosis and mitochondrial and structural damage.
Keyword
MeSH Terms
Figure
-

Fig. 1. Nicotine-induced embryotoxicity at 72 hours post-fertilization (hpf). (A) Hatching/non-hatching embryo findings. The number of embryos showing abnormal morphology tended to be increased in the nicotine-exposed groups (arrow: bending of the trunk). All images were captured at 72 hpf (×32). Scale bar=1,000 μm. (B) Hatching rate, mortality rate, teratogenicity rate, and total abnormal findings (mortality rate+teratogenicity rate) showed statistically significant differences among nicotine concentrations (n=100 per concentration, linear by linear association). Nicotine-exposed groups showed a lower hatching rate and higher mortality and teratogenicity than the control group. The rate of teratogenicity and total abnormal findings increased in a dose-dependent manner (1% in the normal control group, 16% for 5 μM nicotine, 26% for 10 μM nicotine, 37% for 20 μM nicotine, and 53% for 40 μM nicotine; (total n=400). *Statistically significant. All data were evaluated at 72 hpf.
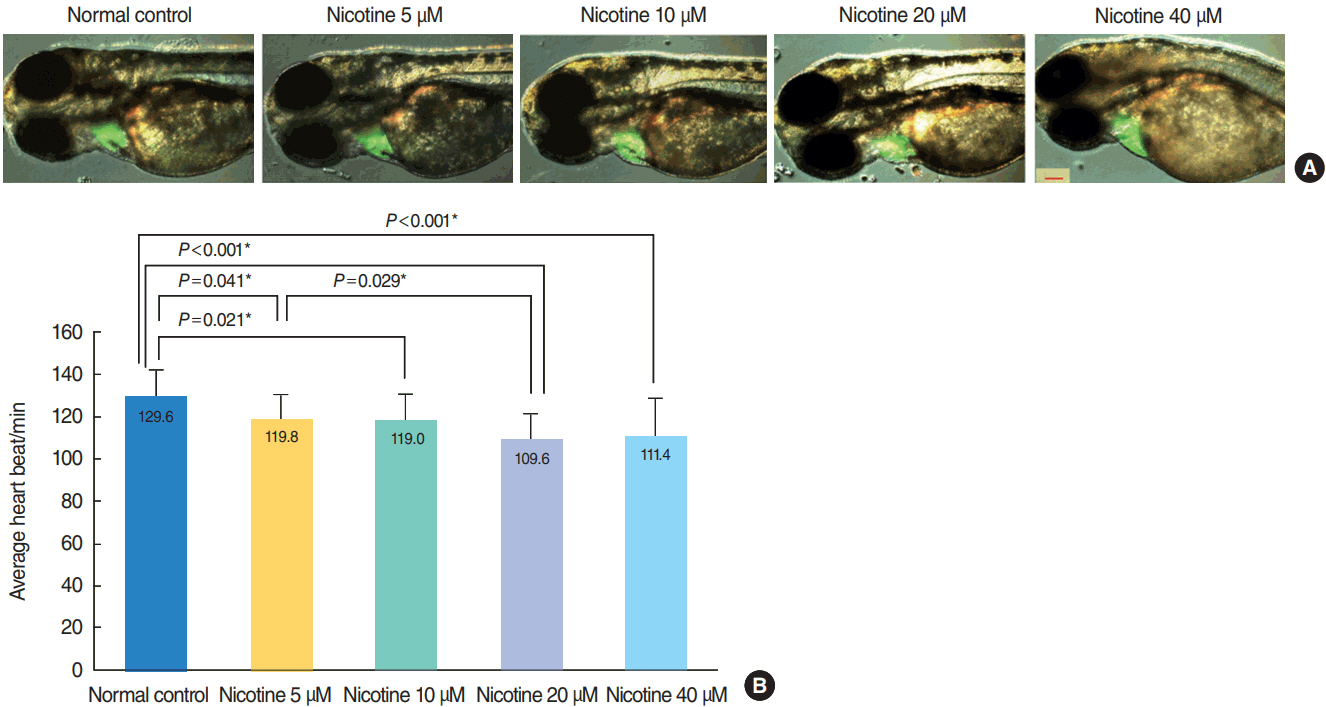
Fig. 2. Heart morphology evaluation and changes in heart rate induced by nicotine treatment. (A) Merged images from the fluorescent image and optical image are shown. There was no difference in heart size between groups (green area). All images were captured using transgenic (cmlc2:EGFP) zebrafish at 72 hours post-fertilization (hpf, ×16). Scale bar=200 μm. (B) The heart rate of zebrafish embryos was significantly decreased in nicotine-exposed groups (P <0.001, one-way analysis of variance). All data were evaluated at 72 hpf. Only statistically significant pair-wise comparisons in post-hoc analysis are shown. *Statistically B significant (P<0.05, total n=150).
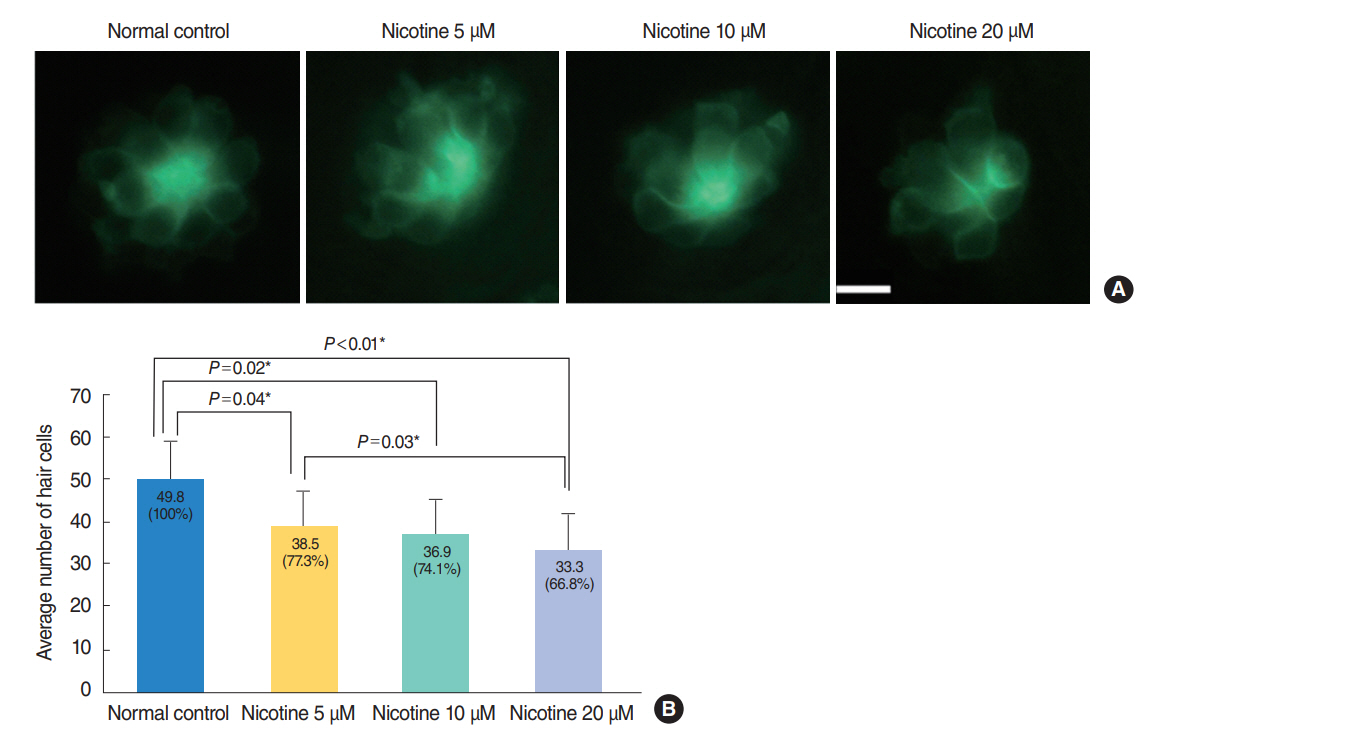
Fig. 3. Nicotine-induced hair cell toxicity at 120 hours post-fertilization (hpf). The total number of hair cells was measured in four neuromasts (supraorbital [SO1 and SO2], otic [O1], and occipital [OC1]; total n=150). (A) The number of hair cells decreased in nicotine-exposed groups as shown by fluorescent microscopy (OC1, ×40). Scale bar=10 μm (×10). (B) Animals in the normal control group had an average of 49.8 hair cells. Nicotine exposure significantly decreased the number of total hair cells of the four neuromasts compared with that of the normal control, (P<0.001, one-way analysis of variance). At a concentration of 40 μM nicotine almost all the fish were dead, therefore statistical analysis was unavailable. All data were evaluated using transgenic (brn3c:EGFP) zebrafish at 120 hpf. Only statistically significant pair-wise comparisons in posthoc analysis are shown. *Statistically significant (P<0.05).

Fig. 4. Nicotine-induced apoptosis in hair cells within neuromasts. Nicotine-induced apoptosis was confirmed by terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling (TUNEL) assay. (A) Green dots represent hair cells. Apoptotic cells appear as light red dots in the merged image obtained by fluorescent microscopy after the TUNEL reaction. All images were obtained using transgenic (brn3c:EGFP) zebrafish at 120 hours post-fertilization. Scale bar=10 μm (×40). (B) The average number of TUNEL-positive cells was measured in each neuromast (supraorbital [SO1, SO2], otic [O1], and occipital [OC1]; total n=50). At a concentration of 40 μM, the fish were almost all dead and statistical analysis was unavailable. The number of TUNEL-positive cells was significantly increased in nicotine-exposed groups (P<0.001, one-way analysis of variance). Only statistically significant pair-wise comparisons in post-hoc analysis are shown. *Statistically significant (P<0.05).
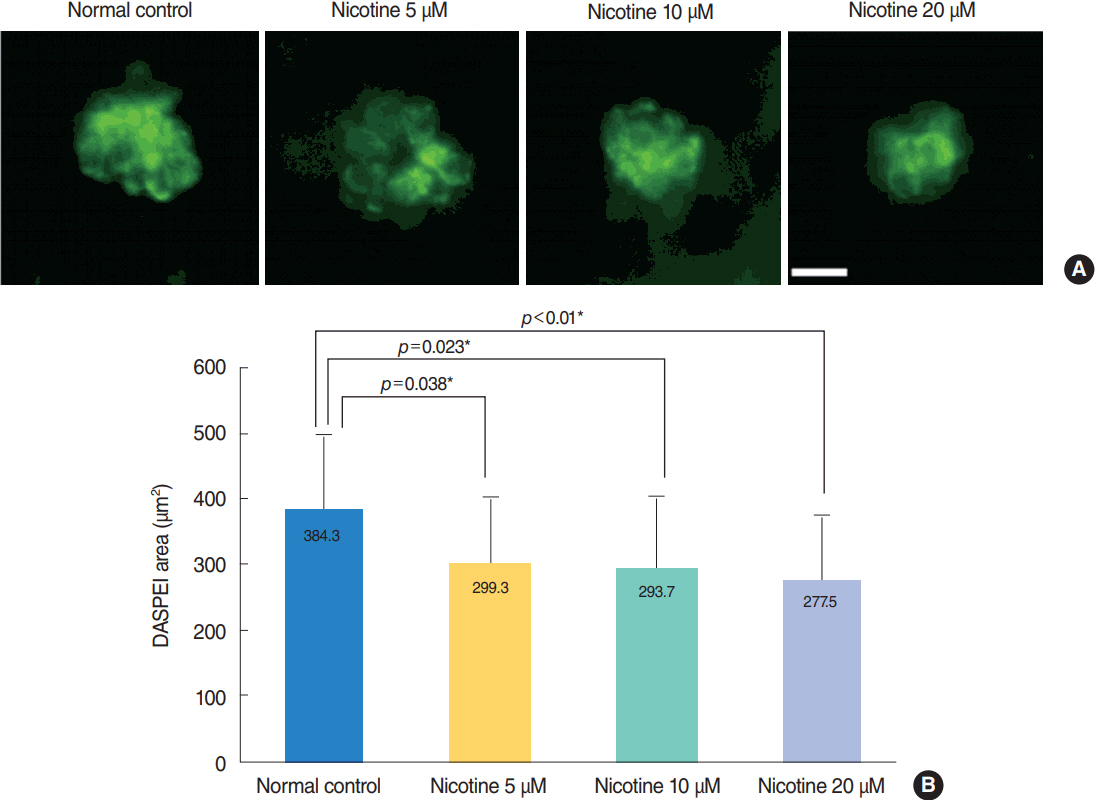
Fig. 5. Analysis of hair cell damage by 2-(4-[dimethylamino]styryl)-N-ethylpyridinium iodide (DASPEI) assay. (A) Staining of hair cells with DASPEI showed that treatment with nicotine decreased the number of hair cells in neuromasts. Scale bar=10 μm (×40). (B) Average DASPEIstained area in four neuromasts (supraorbital [SO1, SO2], otic [O1], and occipital [OC1]; total n=30) significantly decreased as nicotine concentration increased (P<0.001, one-way analysis of variance). Fish exposed to 40 μM nicotine were almost all dead after 120 hours post-fertilization. Only statistically significant pair-wise comparisons in post-hoc analysis are shown. *Statistically significant (P<0.05).

Fig. 6. Reactive oxygen species (ROS) production by 2,7-dichlorodihydrofluorescein diacetate (DCHF-DA) assay. For evaluation of ROS generation, wild type zebrafish was used with DCHF-DA assay on hair cells as in green dots in the middle row. A 4´,6-diamidino-2-phenylindole (DAPI)-positive cells were observed as blue fluorescent dots as in the lower row. By merging those figures, the ROS production cells could be clearly identified as in the upper row. Neomycin as positive control showed strongest ROS generation. Despite not strong as in neomycin, there was increased ROS generation in hair cells in nicotine 40 μM. O1, otic.
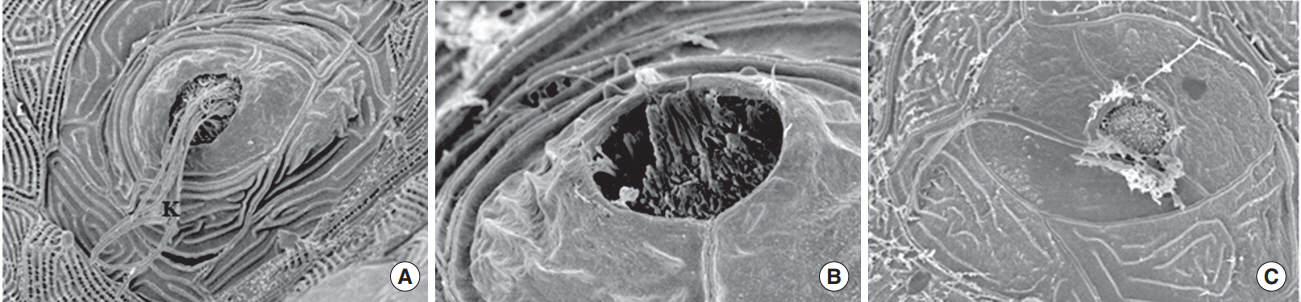
Fig. 7. Scanning electron microscopy. When transgenic (brn3c:EGFP) zebrafish were exposed to nicotine for 120 hours, the kinocilia of hair cells in neuromasts were clearly observed in the normal control (A) and 5 μM nicotine group (B), but not in the 20 μM nicotine group (C). All images were obtained in three zebrafish at 120 hours post-fertilization for each group. Scale bar (at the bottom of each figure, one space)=5 or 10 µm.
Reference
-
1. Gray NJ. Nicotine yesterday, today, and tomorrow: a global review. Nicotine Tob Res. 2014; Feb. 16(2):128–36.
Article2. Abbott LC, Winzer-Serhan UH. Smoking during pregnancy: lessons learned from epidemiological studies and experimental studies using animal models. Crit Rev Toxicol. 2012; Apr. 42(4):279–303.
Article3. Parker B, Connaughton VP. Effects of nicotine on growth and development in larval zebrafish. Zebrafish. 2007; Spring. 4(1):59–68.
Article4. Yoo MH, Rah YC, Choi J, Park S, Park HC, Oh KH, et al. Embryotoxicity and hair cell toxicity of silver nanoparticles in zebrafish embryos. Int J Pediatr Otorhinolaryngol. 2016; Apr. 83:168–74.
Article5. Klee EW, Ebbert JO, Schneider H, Hurt RD, Ekker SC. Zebrafish for the study of the biological effects of nicotine. Nicotine Tob Res. 2011; May. 13(5):301–12.
Article6. Zhao S, Cui WY, Cao J, Luo C, Fan L, Li MD. Impact of maternal nicotine exposure on expression of myelin-related genes in zebrafish larvae. Zebrafish. 2014; Feb. 11(1):10–6.
Article7. Lantz-McPeak S, Guo X, Cuevas E, Dumas M, Newport GD, Ali SF, et al. Developmental toxicity assay using high content screening of zebrafish embryos. J Appl Toxicol. 2015; Mar. 35(3):261–72.
Article8. Ellis LD, Soo EC, Achenbach JC, Morash MG, Soanes KH. Use of the zebrafish larvae as a model to study cigarette smoke condensate toxicity. PLoS One. 2014; Dec. 9(12):e115305.
Article9. Palpant NJ, Hofsteen P, Pabon L, Reinecke H, Murry CE. Cardiac development in zebrafish and human embryonic stem cells is inhibited by exposure to tobacco cigarettes and e-cigarettes. PLoS One. 2015; May. 10(5):e0126259.
Article10. Welsh L, Tanguay RL, Svoboda KR. Uncoupling nicotine mediated motoneuron axonal pathfinding errors and muscle degeneration in zebrafish. Toxicol Appl Pharmacol. 2009; May. 237(1):29–40.
Article11. Chang J, Ryou N, Jun HJ, Hwang SY, Song JJ, Chae SW. Effect of cigarette smoking and passive smoking on hearing impairment: data from a population-based study. PLoS One. 2016; Jan. 11(1):e0146608.
Article12. Dawes P, Cruickshanks KJ, Moore DR, Edmondson-Jones M, McCormack A, Fortnum H, et al. Cigarette smoking, passive smoking, alcohol consumption, and hearing loss. J Assoc Res Otolaryngol. 2014; Aug. 15(4):663–74.
Article13. Paschoal CP, Azevedo MF. Cigarette smoking as a risk factor for auditory problems. Braz J Otorhinolaryngol. 2009; Nov-Dec. 75(6):893–902.
Article14. Durante AS, Pucci B, Gudayol N, Massa B, Gameiro M, Lopes C. Tobacco smoke exposure during childhood: effect on cochlear physiology. Int J Environ Res Public Health. 2013; Oct. 10(11):5257–65.
Article15. Durante AS, Ibidi SM, Lotufo JP, Carvallo RM. Maternal smoking during pregnancy: impact on otoacoustic emissions in neonates. Int J Pediatr Otorhinolaryngol. 2011; Sep. 75(9):1093–8.
Article16. Sun W, Hansen A, Zhang L, Lu J, Stolzberg D, Kraus KS. Neonatal nicotine exposure impairs development of auditory temporal processing. Hear Res. 2008; Nov. 245(1-2):58–64.
Article17. Talaat HS, Metwaly MA, Khafagy AH, Abdelraouf HR. Dose passive smoking induce sensorineural hearing loss in children? Int J Pediatr Otorhinolaryngol. 2014; Jan. 78(1):46–9.18. Abdel-Hafez AM, Elgayar SA, Husain OA, Thabet HS. Effect of nicotine on the structure of cochlea of guinea pigs. Anat Cell Biol. 2014; Sep. 47(3):162–70.
Article19. Rah YC, Choi J, Yoo MH, Yum G, Park S, Oh KH, et al. Ecabet sodium alleviates neomycin-induced hair cell damage. Free Radic Biol Med. 2015; Dec. 89:1176–83.
Article20. Chang J, Choi J, Rah YC, Yoo MH, Oh KH, Im GJ, et al. Sodium selenite acts as an otoprotectant against neomycin-induced hair cell damage in a zebrafish model. PLoS One. 2016; Mar. 11(3):e0151557.
Article21. Kakehata S, Nakagawa T, Takasaka T, Akaike N. Cellular mechanism of acetylcholine-induced response in dissociated outer hair cells of guinea-pig cochlea. J Physiol. 1993; Apr. 463:227–44.
Article22. Matta SG, Balfour DJ, Benowitz NL, Boyd RT, Buccafusco JJ, Caggiula AR, et al. Guidelines on nicotine dose selection for in vivo research. Psychopharmacology (Berl). 2007; Feb. 190(3):269–319.
Article23. Ackerman KM, Nakkula R, Zirger JM, Beattie CE, Boyd RT. Cloning and spatiotemporal expression of zebrafish neuronal nicotinic acetylcholine receptor alpha 6 and alpha 4 subunit RNAs. Dev Dyn. 2009; Apr. 238(4):980–92.
Article24. Thomas LT, Welsh L, Galvez F, Svoboda KR. Acute nicotine exposure and modulation of a spinal motor circuit in embryonic zebrafish. Toxicol Appl Pharmacol. 2009; Aug. 239(1):1–12.
Article25. Svoboda KR, Vijayaraghavan S, Tanguay RL. Nicotinic receptors mediate changes in spinal motoneuron development and axonal pathfinding in embryonic zebrafish exposed to nicotine. J Neurosci. 2002; Dec. 22(24):10731–41.
Article26. Rao P, Ande A, Sinha N, Kumar A, Kumar S. Effects of cigarette smoke condensate on oxidative stress, apoptotic cell death, and HIV replication in human monocytic cells. PLoS One. 2016; May. 11(5):e0155791.
Article27. Nathiga Nambi KS, Abdul Majeed S, Taju G, Sivasubbu S, Sarath Babu V, Sahul Hameed AS. Effects of nicotine on zebrafish: a comparative response between a newly established gill cell line and whole gills. Comp Biochem Physiol C Toxicol Pharmacol. 2017; May. 195:68–77.
Article28. Fabry DA, Davila EP, Arheart KL, Serdar B, Dietz NA, Bandiera FC, et al. Secondhand smoke exposure and the risk of hearing loss. Tob Control. 2011; Jan. 20(1):82–5.
Article29. Mustafa MW. Transient evoked otoacoustic emissions and vestibular evoked myogenic potentials in cigarette and water pipe smokers. Eur Arch Otorhinolaryngol. 2014; Oct. 271(10):2669–73.
Article30. Weitzman M, Govil N, Liu YH, Lalwani AK. Maternal prenatal smoking and hearing loss among adolescents. JAMA Otolaryngol Head Neck Surg. 2013; Jul. 139(7):669–77.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- microRNA-183 is Essential for Hair Cell Regeneration after Neomycin Injury in Zebrafish
- Effect of Antioxidant and NMDA Receptor Antagonist on Mercury-Induced Toxicity in Cultured Preimplantation Embryos
- Zebrafish and Mycobacterial infection
- Experimental Studies on the Relation between Nicotine and Sexual Hormone: Part IV. The Antidotal Action of Luteohormone on Nicotine Toxicity during Anaphylaxis: C. The Effect of Nicotine with Luteohormone on Anaphylaxis
- Role of histone deacetylase activity in the developing lateral line neuromast of zebrafish larvae
