Potassium Currents in Isolated Deiters' Cells of Guinea Pig
- Affiliations
-
- 1Department of Otolaryngology, Asan Medical Center and University of Ulsan College of Medicine, Seoul 138-736, Korea.
- 2Department of Otolaryngology, Kangwon National University College of Medicine, Chuncheon 200-701, Korea.
- 3Department of Physiology, Asan Medical Center and University of Ulsan College of Medicine, Seoul 138-736, Korea. leemch@amc.seoul.kr
- 4Department of Physiology, College of Medicine, Cardiovascular and Metabolic Disease Center, Inje University, Busan 614-735, Korea.
- KMID: 2285482
- DOI: http://doi.org/10.4196/kjpp.2013.17.6.537
Abstract
- Deiters' cells are the supporting cells in organ of Corti and are suggested to play an important role in biochemical and mechanical modulation of outer hair cells. We successfully isolated functionally different K+ currents from Deiters' cells of guinea pig using whole cell patch clamp technique. With high K+ pipette solution, depolarizing step pulses activated strongly outward rectifying currents which were dose-dependently blocked by clofilium, a class III anti-arrhythmic K+ channel blocker. The remaining outward current was transient in time course whereas the clofilium-sensitive outward current showed slow inactivation and delayed rectification. Addition of 5 mM tetraethylammonium (TEA) further blocked the remaining current leaving a very fast inactivating transient outward current. Therefore, at least three different types of K+ current were identified in Deiters' cells, such as fast activating and fast inactivating current, fast activating slow inactivating current, and very fast inactivating transient outward current. Physiological role of them needs to be established.
MeSH Terms
Figure
-

Fig. 1 Isolated Deiters' cell. The cell is easily recognized by its long phalangeal process (arrow) and nucleus in cell body.
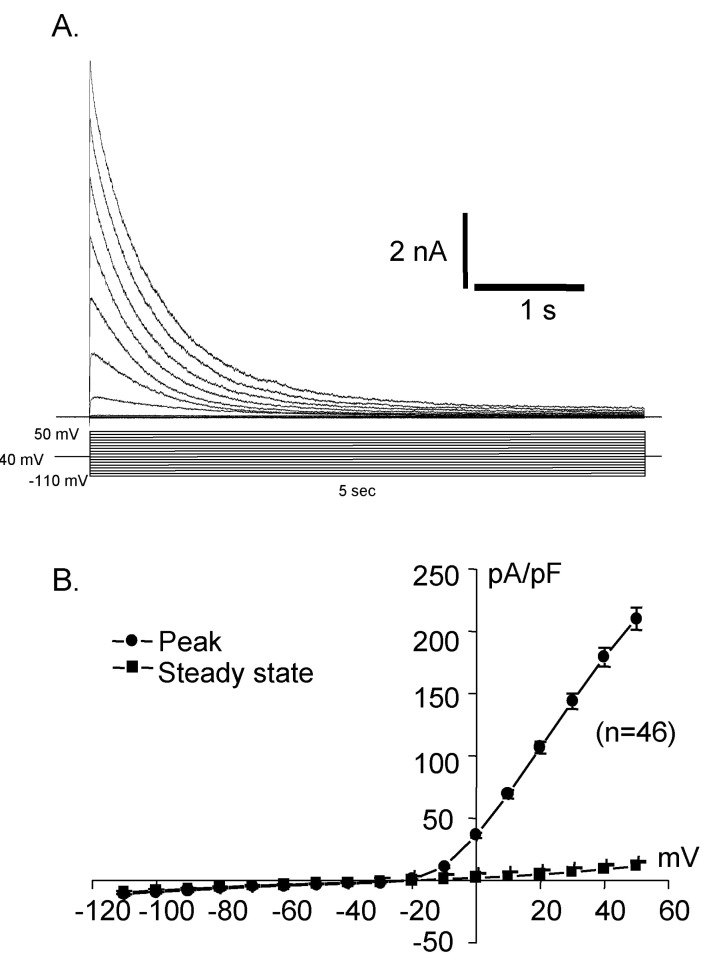
Fig. 2 Activated current traces (A) and I-V curve (B) recorded from Deiters' cells. This shows rapid activating outward rectifying current. Holding at -40 mV in whole cell recording, step pulse with 10 mV increment from -110 mV to 50 mV elicited outward current, showing very strong outward rectification.

Fig. 3 Effects of extracellular calcium change on the activated current traces and their I-V curves. (A) 2 mM Ca2+, (B) 0 mM Ca2+, (C) I-V curves. 2 mM Ca2+ (●) and 0 mM Ca2+ (■).
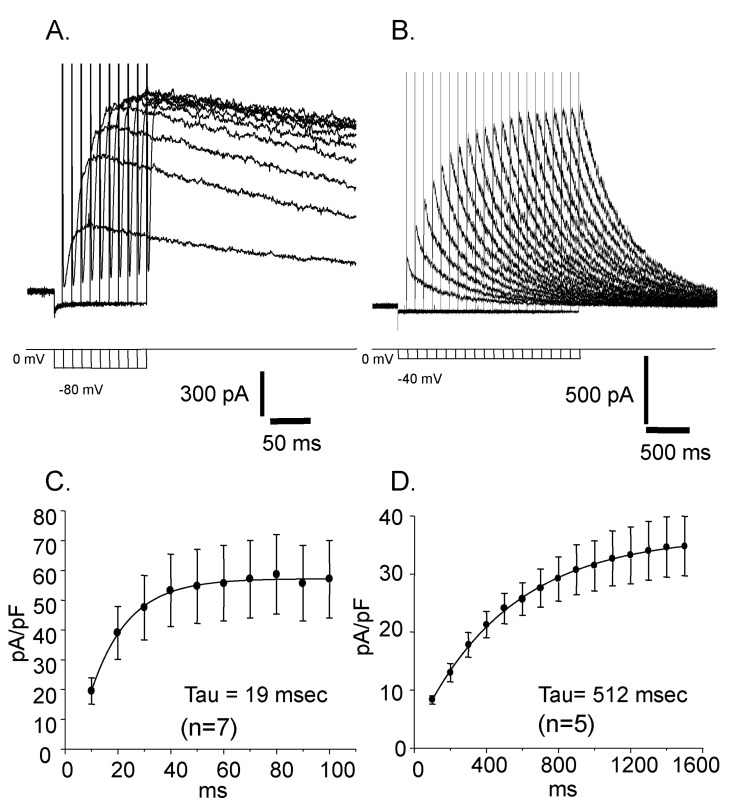
Fig. 4 Voltage dependent changes of the reactivation time courses. (A) The voltage of prepulse was -80 mV. (B) The voltage of prepulse was -40 mV. (C) The time constant of the current reactivation was 19 msec. (D) The time constant of the current reactivation was 512 msec.
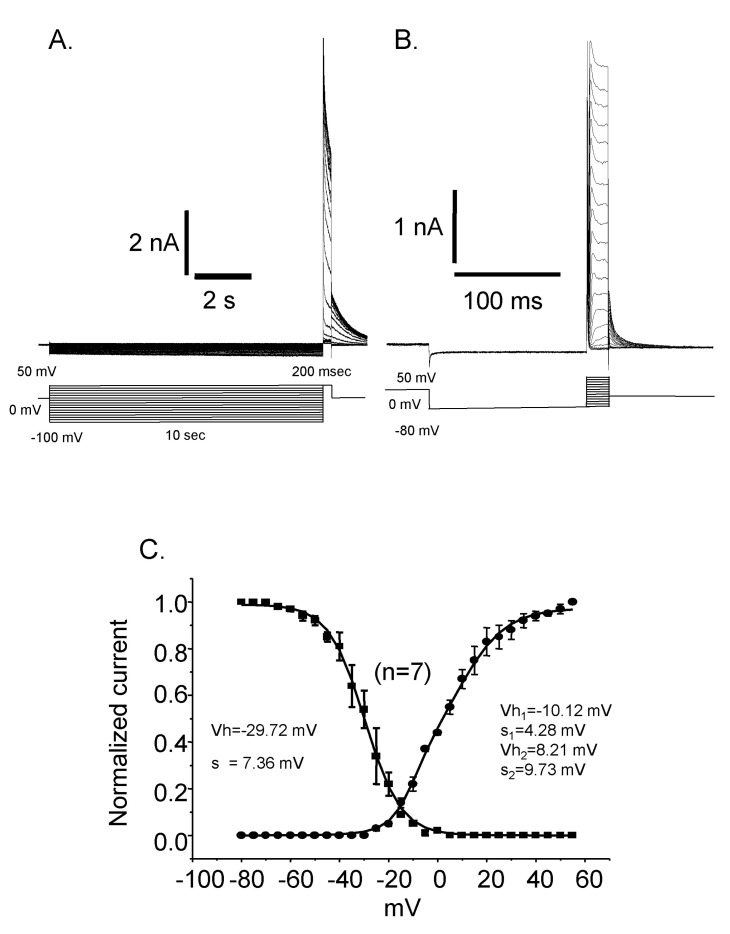
Fig. 5 Steady state inactivation and activation curves. (A) Current traces of the steady state inactivation. (B) Current traces of the activation. (C) The activation curve is fitted by double Boltzmann equation suggesting the presence of more than two channels. Vh: half activation (or inactivation) voltage, K: slope factor. Inactivation curve could be fitted by single Boltzmann equation and the half inactivation voltage (Vh) was -29.72 mV and the slope factor (s) was 7.36. Activation curve was fitted by double Boltzmann equation. The half activation voltages (Vh) and slope factors (s) were -10.12 mV and 4.28 for the first one and 8.21 mV and 9.73 for the second one, respectively.
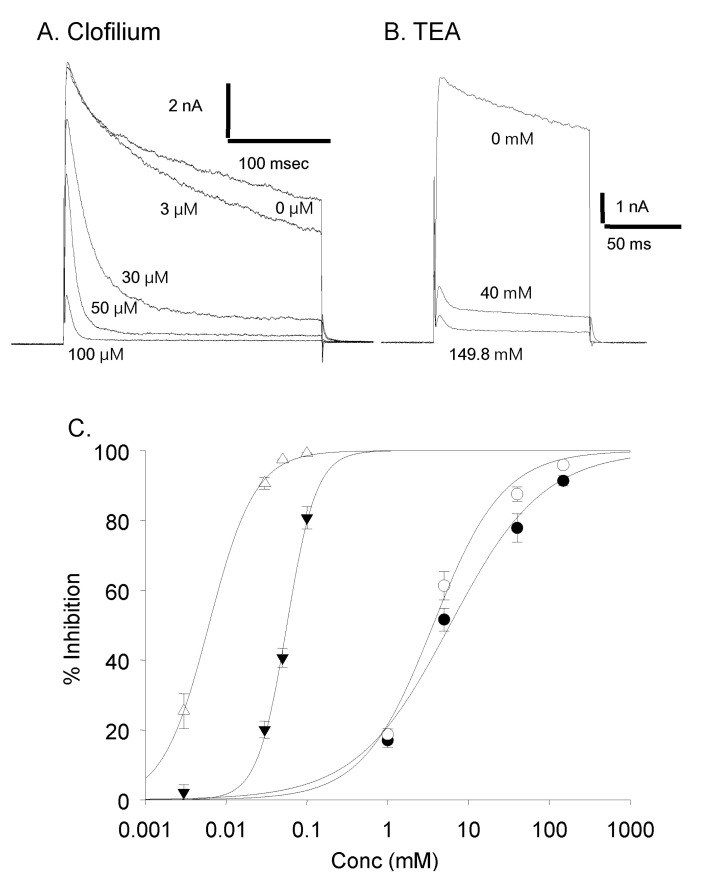
Fig. 6 Dose dependent effect of clofilium and TEA. (A) Current traces with different concentration of clofilium. (B) Current traces with different concentration of TEA. (C) Dose response curves. Effect of TEA on the peak current amplitude (●) and on the current amplitude at 100 msec (◯) (1 mM, n=4; 5 mM, n=11; 40 mM, n=7; 149.8 mM, n=3). Effect of clofilium on the peak current amplitude (▼) and on the current amplitude at 100 msec (△) (3 µM, n=5; 30 µM, n=12; 50 µM, n=5; 100 µM, n=5).
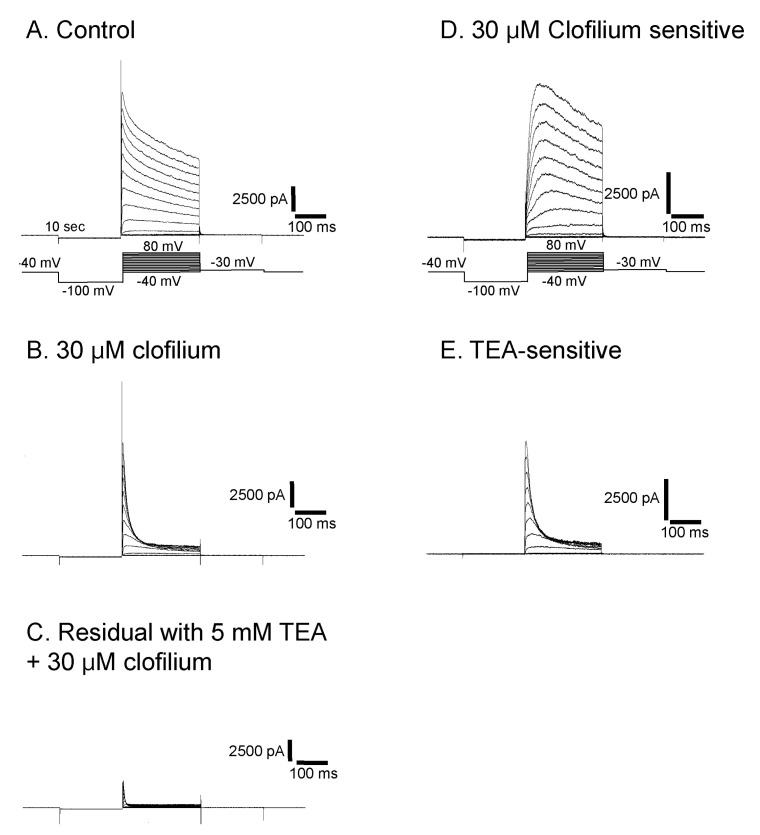
Fig. 7 Isolation of three different types of currents in Deiters' cells. At a concentration of 30 µM of clofilium, control outward rectifying current (A) was blocked leaving transient outward current (B). This residual current was further blocked by 5 mM TEA and only a small transient outward current was identified (C). Clofilium-sensitive current (D) was obtained by subtracting (B) from (A). TEA-sensitive current (E) was obtained by subtracting (C) from (B).
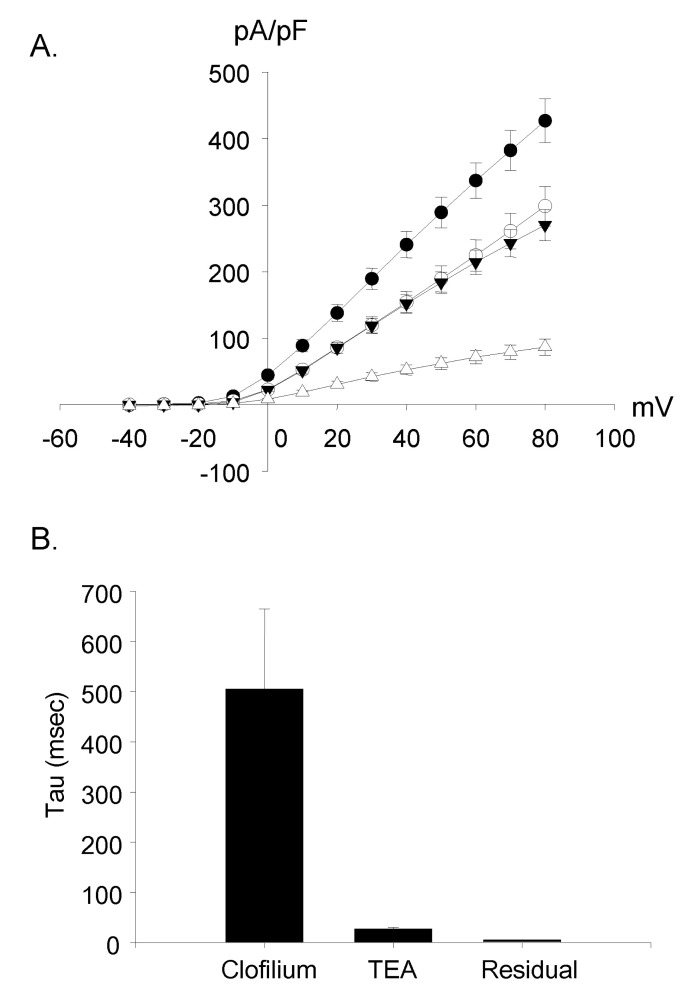
Fig. 8 Voltage-current relationship of three currents. (A) The I-V curves of three currents were similar to that of control currents. The peak current amplitude was used. All the curves showed outward rectification. ●: control current (n=12); ◯: clofiliumsensitive current (n=9); ▼: TEA-sensitive current (n=9); △: clofilium and TEA resistant current (n=9). (B) The time constant (tau) was 505±160 ms in the clofilium-sensitive current, 27.6±3.1 ms in the TEA-sensitive current and 5.6±0.1 ms in the residual current (n=9).
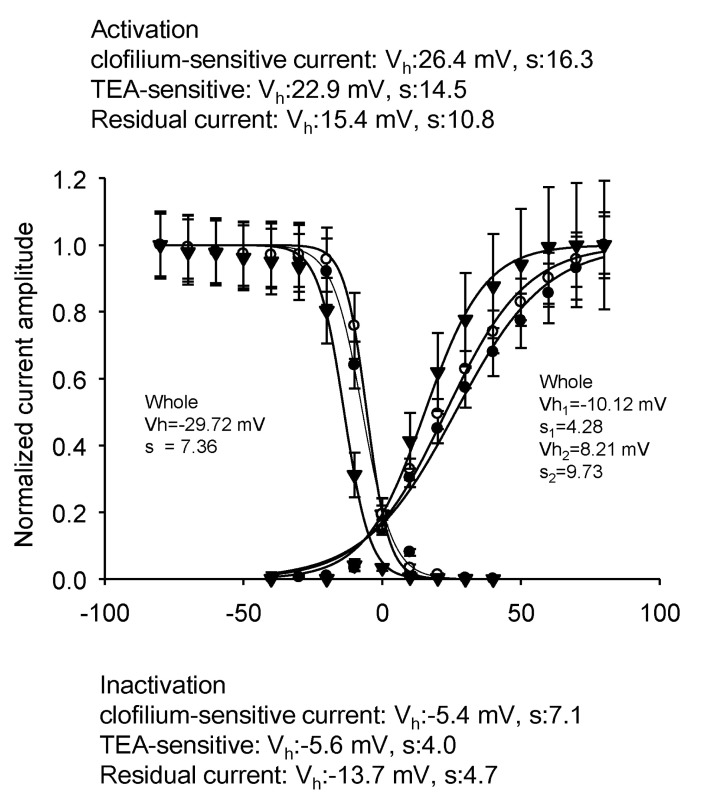
Fig. 9 Characteristics of activation and inactivation of three different currents. the clofilium-sensitive current (●); TEA-sensitive current (◯); clofilium and TEA resistant current (▼). All the currents were fitted by each single Boltzmann equation. Each fitting parameters were shown on the figure.
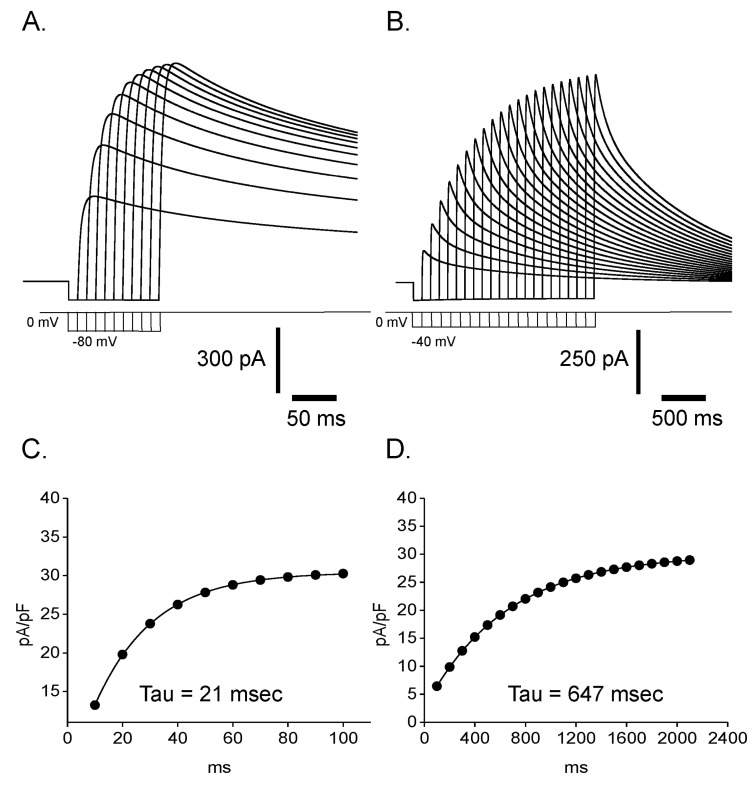
Fig. 10 Validation of our mathematical model. Reactivation time courses from prepulse at -80 mV (A) and -40 mV (B) were obtained by our mathematical simulation. By plotting current density (pA/pF) against length of prepulse and fitting them to single exponential functions (C, D), time constants of reactivation were estimated to be 21 msec at -80 mV and 647 msec at -40 mV, respectively which is comparable to experimentally obtained values in Fig. 4 (19 msec at -80 mV and 512 msec at -40 mV).
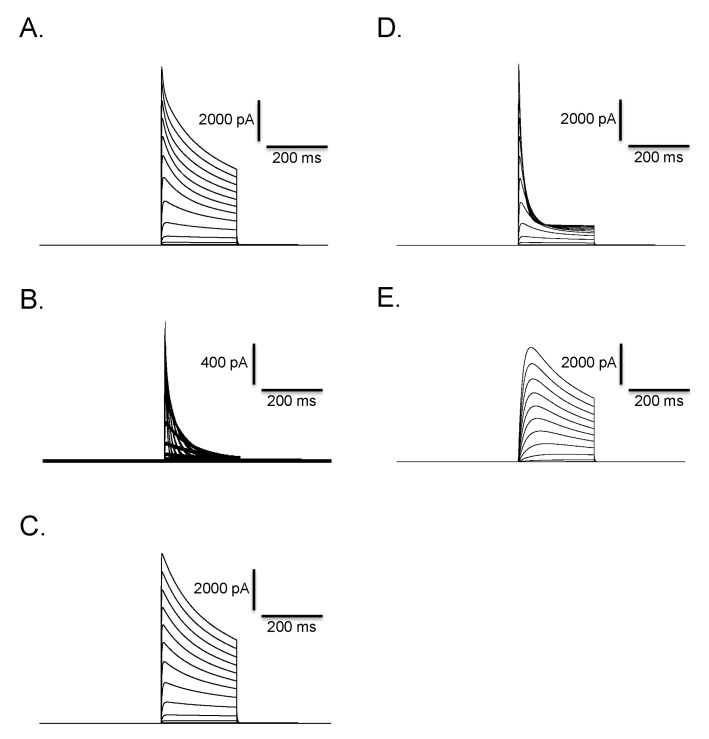
Fig. 11 Mathematical simulation of outward K+ current and clofilium effect in Deiters' cells. Fast activating and fast inactivating current (B) and fast activating and slow inactivating current (C) were summated to reproduce experimentally obtained outward rectifying K+ current in Fig. 7A. Summated current (A) clearly shows biphasic time course of inactivation. The pulse protocol is identical with that used in Fig. 7A. Incorporation of transition from open to blocked state into the C-O-I model successfully reproduced the effect of clofilium in Fig. 7B. Time course and voltage-dependence of simulated current (D) are similar with those of experimentally obtained current in Fig. 7B. Simulated clofilium-sensitive current (E) was obtained by subtracting (D) from (A). Time course and voltage-dependence of simulated current are very similar to those of experimentally obtained current in Fig. 7D.
Reference
-
1. Liberman MC, Dodds LW, Pierce S. Afferent and efferent innervation of the cat cochlea: quantitative analysis with light and electron microscopy. J Comp Neurol. 1990; 301:443–460. PMID: 2262601.
Article2. Wright CG, Preston RE. Efferent nerve fibers associated with the outermost supporting cells of the organ of Corti in the guinea pig. Acta Otolaryngol. 1976; 82:41–47. PMID: 948984.
Article3. Burgess BJ, Adams JC, Nadol JB Jr. Morphologic evidence for innervation of Deiters' and Hensens cells in the guinea pig. Hear Res. 1997; 108:74–82. PMID: 9213124.
Article4. Matsunobu T, Chung JW, Schacht J. Acetylcholine-evoked calcium increases in Deiters' cells of the guinea pig cochlea suggest alpha9-like receptors. J Neurosci Res. 2001; 63:252–256. PMID: 11170174.5. Chung JW, Schacht J. ATP and nitric oxide modulate intracellular calcium in isolated pillar cells of the guinea pig cochlea. J Assoc Res Otolaryngol. 2001; 2:399–407. PMID: 11833612.
Article6. Matsunobu T, Schacht J. Nitric oxide/cyclic GMP pathway attenuates ATP-evoked intracellular calcium increase in supporting cells of the guinea pig cochlea. J Comp Neurol. 2000; 423:452–461. PMID: 10870085.
Article7. Bobbin RP. ATP-induced movement of the stalks of isolated cochlear Deiters' cells. Neuroreport. 2001; 12:2923–2926. PMID: 11588604.
Article8. Dulon D, Blanchet C, Laffon E. Photo-released intracellular Ca2+ evokes reversible mechanical responses in supporting cells of the guinea-pig organ of Corti. Biochem Biophys Res Commun. 1994; 201:1263–1269. PMID: 8024570.9. Fridberger A, Flock A, Ulfendahl M, Flock B. Acoustic overstimulation increases outer hair cell Ca2+ concentrations and causes dynamic contractions of the hearing organ. Proc Natl Acad Sci U S A. 1998; 95:7127–7132. PMID: 9618550.10. Nenov AP, Chen C, Bobbin RP. Outward rectifying potassium currents are the dominant voltage activated currents present in Deiters' cells. Hear Res. 1998; 123:168–182. PMID: 9745965.
Article11. Szucs A, Somodi S, Batta TJ, Tóth A, Szigeti GP, Csernoch L, Panyi G, Sziklai I. Differential expression of potassium currents in Deiters' cells of the guinea pig cochlea. Pflugers Arch. 2006; 452:332–341. PMID: 16447073.12. Qu C, Liang F, Hu W, Shen Z, Spicer SS, Schulte BA. Expression of CLC-K chloride channels in the rat cochlea. Hear Res. 2006; 213:79–87. PMID: 16466872.
Article13. Castle NA. Selective inhibition of potassium currents in rat ventricle by clofilium and its tertiary homolog. J Pharmacol Exp Ther. 1991; 257:342–350. PMID: 2019997.14. Steidl JV, Yool AJ. Distinct mechanisms of block of Kv1.5 channels by tertiary and quaternary amine clofilium compounds. Biophys J. 2001; 81:2606–2613. PMID: 11606274.
Article15. Yeola SW, Snyders DJ. Electrophysiological and pharmacological correspondence between Kv4.2 current and rat cardiac transient outward current. Cardiovasc Res. 1997; 33:540–547. PMID: 9093524.
Article16. Dixon JE, Shi W, Wang HS, McDonald C, Yu H, Wymore RS, Cohen IS, McKinnon D. Role of the Kv4.3 K+ channel in ventricular muscle. A molecular correlate for the transient outward current. Circ Res. 1996; 79:659–668. PMID: 8831489.17. Rudy B, Kentros C, Vela-Saenz De Miera E. Families of potassium channel genes in mammals: Toward an understanding of the molecular basis of potassium channel diversity. Mol Cell Neurosci. 1991; 2:89–102. PMID: 19912787.
Article18. Shieh CC, Coghlan M, Sullivan JP, Gopalakrishnan M. Potassium channels: molecular defects, diseases, and therapeutic opportunities. Pharmacol Rev. 2000; 52:557–594. PMID: 11121510.19. Wangemann P. K+ cycling and the endocochlear potential. Hear Res. 2002; 165:1–9. PMID: 12031509.20. Marcus DC, Wu T, Wangemann P, Kofuji P. KCNJ10 (Kir4.1) potassium channel knockout abolishes endocochlear potential. Am J Physiol Cell Physiol. 2002; 282:C403–C407. PMID: 11788352.
Article21. Mori N, Sakagami M, Fukazawa K, Matsunaga T. An immunohistochemical and electrophysiological study on Isk protein in the stria vascularis of the guinea pig. Eur Arch Otorhinolaryngol. 1993; 250:186–189. PMID: 8357611.
Article22. Rossi ML, Ferrary E, Martini A, Martini M, Pelucchi B, Bernard C, Teixeira M, Sterkers O, Fesce R. The effect of clofilium, a K-channel blocker, on the electrogenic K secretion and the sensory discharge at the frog semicircular canal. Brain Res. 1996; 721:174–180. PMID: 8793098.
Article23. Lagostena L, Cicuttin A, Inda J, Kachar B, Mammano F. Frequency dependence of electrical coupling in Deiters' cells of the guinea pig cochlea. Cell Commun Adhes. 2001; 8:393–399. PMID: 12064625.
Article24. Yang J, Wang J. Possible function of outward potassium currents in isolated Deiters' cells of guinea pig cochlea. Chin Med J (Engl). 2002; 115:264–267. PMID: 11940345.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Outward Rectifying Current in Isolated Deiters' Cells from Guinea Pig Cochlea
- Temperature Enhances Activation and Inactivation Kinetics of Potassium Currents in Inner Hair Cells Isolated from Guinea-Pig Cochlea
- Modulation of outward potassium currents by nitric oxide in longitudinal smooth muscle cells of guinea-pig ileum
- Effects of Aminoglycoside Antibiotics on Acetylcholine-induced Potassium Currents in Guinea-pig Outer Hair Cell
- The Effects of Neurotransmitters on the Ion Channels in the Isolated Deiters'Cells of Guinea Pig Cochlea
