Angiotensin II and TGF-β1 Induce Alterations in Human Amniotic Fluid-Derived Mesenchymal Stem Cells Leading to Cardiomyogenic Differentiation Initiation
- Affiliations
-
- 1Department of Molecular Cell Biology, Institute of Biochemistry, Life Sciences Center, Vilnius University, Vilnius, Lithuania. monika.gasiuniene@gmc.vu.lt
- 2Electronic Systems Department, Electronics Faculty, Vilnius Gediminas Technical University, Vilnius, Lithuania.
- KMID: 2465896
- DOI: http://doi.org/10.15283/ijsc18126
Abstract
- BACKGROUND AND OBJECTIVES
Human amniotic fluid-derived mesenchymal stem cells (AF-MSCs) may be a valuable source for cardiovascular tissue engineering and cell therapy. The aim of this study is to verify angiotensin II and transforming growth factor-beta 1 (TGF-β1) as potential cardiomyogenic differentiation inducers of AF-MSCs.
METHODS AND RESULTS
AF-MSCs were obtained from amniocentesis samples from second-trimester pregnant women, isolated and characterized by the expression of cell surface markers (CD44, CD90, CD105 positive; CD34 negative) and pluripotency genes (OCT4, SOX2, NANOG, REX1). Cardiomyogenic differentiation was induced using different concentrations of angiotensin II and TGF-β1. Successful initiation of differentiation was confirmed by alterations in cell morphology, upregulation of cardiac genes-markers NKX2-5, TBX5, GATA4, MYH6, TNNT2, DES and main cardiac ion channels genes (sodium, calcium, potassium) as determined by RT-qPCR. Western blot and immunofluorescence analysis revealed the increased expression of Connexin43, the main component of gap junctions, and Nkx2.5, the early cardiac transcription factor. Induced AF-MSCs switched their phenotype towards more energetic and started utilizing oxidative phosphorylation more than glycolysis for energy production as assessed using Agilent Seahorse XF analyzer. The immune analysis of chromatin-modifying enzymes DNMT1, HDAC1/2 and Polycomb repressive complex 1 and 2 (PRC1/2) proteins BMI1, EZH2 and SUZ12 as well as of modified histones H3 and H4 indicated global chromatin remodeling during the induced differentiation.
CONCLUSIONS
Angiotensin II and TGF-β1 are efficient cardiomyogenic inducers of human AF-MSCs; they initiate alterations at the gene and protein expression, metabolic and epigenetic levels in stem cells leading towards cardiomyocyte-like phenotype formation.
Keyword
MeSH Terms
-
Amniocentesis
Amniotic Fluid
Angiotensin II*
Angiotensins*
Blotting, Western
Calcium
Cell Differentiation
Cell- and Tissue-Based Therapy
Chromatin
Chromatin Assembly and Disassembly
Connexin 43
Epigenomics
Female
Fluorescent Antibody Technique
Gap Junctions
Glycolysis
Histones
Humans*
Ion Channels
Mesenchymal Stromal Cells*
Muscle Cells
Oxidative Phosphorylation
Phenotype
Polycomb Repressive Complex 1
Pregnant Women
Smegmamorpha
Stem Cells
Tissue Engineering
Transcription Factors
Up-Regulation
Angiotensin II
Angiotensins
Calcium
Chromatin
Connexin 43
Histones
Ion Channels
Polycomb Repressive Complex 1
Transcription Factors
Figure
-
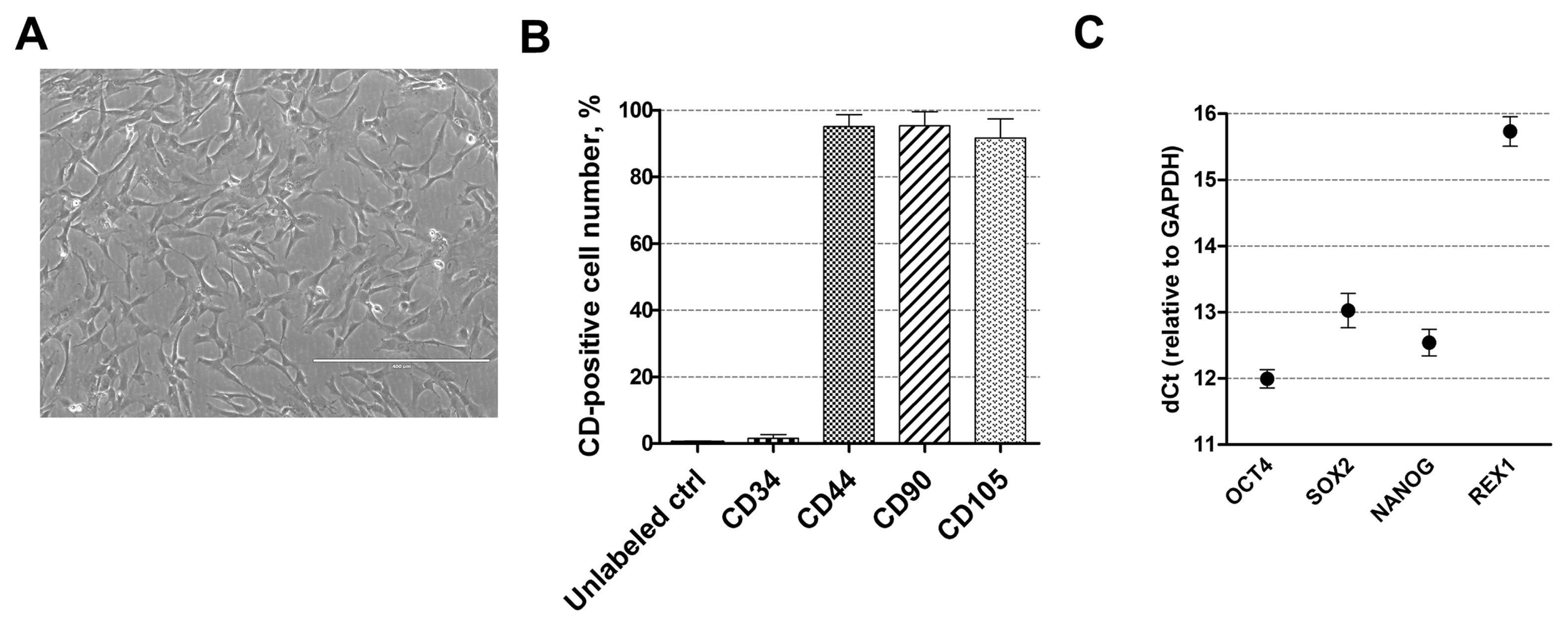
Fig. 1 Characterization of human AF-MSCs. (A) The typical spindle-shaped morphology of human amniotic fluid-derived mesenchymal stem cells, cultivated in cell culture. Scale bar=400 μm. (B) The expression of cell surface markers CD44, CD90, CD105 and CD34 as detected by flow cytometry. Unlabeled ctrl–non-labeled, undifferentiated control cells. Results are presented as mean±SD (n=3). (C) The relative expression of pluripotency genes-markers, such as OCT4, SOX2, NANOG and REX1 as determined by RT-qPCR. Data, relative to GAPDH, are presented as mean±SD (n=3).

Fig. 2 Alterations in morphology and gene expression patterns during decitabine, angiotensin II and TGF-β1 induced cardiac differentiation. (A) Assessment of the cell morphology 12 days after the cardiac differentiation initiation. Dec: decitabine, AngII: angiotensin II, TGF-β1: transforming growth factor-beta 1. Scale bar=400 μm. (B) The connections between proteins that are considered as cardiomyogenic differentiation markers. NKX2-5, TBX5 and GATA4 are the early transcription markers, initiating the expression of structural proteins of cardiomyocytes, such as MYH6 (α-myosin heavy chain), TNNT2 (cardiac troponin T) and DES (Desmin). The data was obtained from the STRING database. (C) The relative expression of the main cardiac : genes-markers, such as NKX2-5, TBX5, GATA4, MYH6, TNNT2 and DES at the days 5 (5 d.) and 12 (12 d.) of induced differentiation. Ctrl–non-differentiated control cells. (D) The relative expression of cardiac ion channels genes at the day 12 of differentiation: SCN5A (sodium voltage-gated channel α-subunit 5), CACNA1D (L-type calcium channel), KCNJ12 and KCND3 (voltage-gated potassium) and HCN2 (hyperpolarization-activated cyclic nucleotide-gated channel). The relative gene expression was determined by RT-qPCR, normalized to GAPDH and presented as a fold change over undifferentiated control. The data are presented as mean±SD (n=3), p≤0.05 (*), p≤0.01 (**), p≤0.001 (***), ns: non-significant.

Fig. 3 Evaluation of Connexin43 (Cx43) and Nkx2.5 levels in induced AF-MSCs. (A) The levels of Cx43, the main gap junction protein, and Nkx2.5, an early cardiac transcription factor, as determined by Western blot in non-differentiated cells (Ctrl) and differentiated cells at the days 5 and 12. As a positive control mouse heart lysate was used. The relative density of each band was measured using ImageJ software (NIH, USA), normalized to the GAPDH loading control and presented as a fold difference over control. The data are presented as mean±SD (n=3), p≤0.05 (*), p≤0.01 (**), p≤0.001 (***), ns: non-significant. The blots represent one of three independent experiments showing similar results. (B) The distribution of Cx43 in control and induced cells at the 12 day of differentiation as obtained using the immunofluorescence assay. The data from each differentiation sample is presented in two images: the left panel–two channels view (blue nuclei and green Cx43) and the right panel–the same image in three channels view (blue nuclei, green Cx43 and red F-actin). Samples were observed using Zeiss Axio Observer fluorescence microscope, 63X magnification with immersion oil, scale bar=10 μm. (C) The immunofluorescence data of Nkx2.5 protein in control and induced cells at the 12 day of differentiation. Nuclei stained blue, Nkx2.5 – red and F-actin – green. Samples were observed using Zeiss Axio Observer fluorescence microscope, 63X magnification with immersion oil, scale bar=10 μm. (D) Quantitative evaluation of Nkx2.5 nuclear localization at the day 12 of differentiation as calculated from the Nkx2.5 immunofluorescence images. The nuclear quantity of Nkx2.5 was normalized to the median value of undifferentiated Control (Ctrl) and presented as normalized nuclear quantity (NNQ). All NNQs were graphed as Tukey-style box plots with sample size n=66, 31, 57, 58, 64 and 60. Relative change of medians and the statistical significance of changes are presented above the box plots. Wilcoxon rank sum test was used for statistical analysis, p≤0.001 (***).

Fig. 4 Cell energy phenotype and metabolic alterations during cardiac differentiation of AF-MSCs. (A) Cell energy diagram showing normalized oxygen consumption rate (OCR) and normalized extracellular acidification rate (ECAR) of control and differentiated cells at the basal (the hollow points) and stressed conditions (colored points). (B) The ratio of normalized OCR to normalized ECAR in control and induced AF-MSCs. (C) Metabolic potential (in percent) of undifferentiated and induced AF-MSCs was calculated from stressed OCR/ECAR over Baseline OCR/ECAR. (D) Mitochondrial respiration expressed as normalized OCR of control and differentiated cells after the addition of ETC inhibitors oligomycin, FCCP, rotenone and antimycin A. The time in the x-axis represents time points when each measurement was done. All calculations of mitochondrial function were done referring to these measurements. (E) Maximal respiration, calculated as normalized OCR fold induction over baseline. (F) Spare respiratory capacity, a derivative from Maximal respiration, was expressed in percent for the control and induced cells. (G) ATP production was calculated as the OCR difference between the baseline and after the addition of oligomycin. (H) The relative expression of genes, related to the cell metabolism and respiration: PPARGC1A (Peroxisome Proliferator-Activated Receptor Gamma Coactivator 1 Alpha), NRF1 (nuclear respiratory factor 1) and HIF1A (hypoxia-inducible factor 1-alpha), in control and differentiated cells as determined using RT-qPCR. The data were normalized to GAPDH and presented as a fold change over undifferentiated control. The data in A~G was obtained using Seahorse XFp Extracellular Flux Analyzer (Agilent, USA), “Cell energy phenotype test” (parts A~C) or “Cell Mito Stress Test” kits (parts D~G) and cells differentiated for 12 days. Results are presented as mean±SD. p≤0.05 (*), p≤0.01 (**), p≤0.001 (***), ns: non-significant.

Fig. 5 Epigenetic alterations during induced cardiomyogenic differentiation. (A) Total proteins were extracted from control cells (Ctrl) and cells, induced using angiotensin II and TGF-β1 at the days 5 and 12 of differentiation. Alterations in the expression levels of Polycomb repressive complex 2 (PRC2) proteins EZH2 and SUZ12, Polycomb repressive complex 1 (PRC1) component BMI1, DNA and histones modifying proteins DNMT1, HDAC1 and HDAC2 as well as in histone modifications, considered as the markers of active chromatin state (H3K4me3, H3K9Ac and H4hyperAc) are presented as determined by Western blot. Mouse heart lysate was used as a positive control. The blots represent one of three independent experiments showing similar results. (B) Relative band density of chromatin remodeling proteins and (C) histone modifications, as measured using ImageJ software (NIH, USA) and normalized to the GAPDH loading control. The data are presented as mean±SD (n=3), p≤0.05 (*), p≤0.01 (**), p≤0.001 (***), ns: non-significant.
Reference
-
References
1. Bagno L, Hatzistergos KE, Balkan W, Hare JM. Mesenchymal stem cell-based therapy for cardiovascular disease: progress and challenges. Mol Ther. 2018; 26:1610–1623. DOI: 10.1016/j.ymthe.2018.05.009. PMID: 29807782. PMCID: 6037203.
Article2. Savickiene J, Treigyte G, Baronaite S, Valiuliene G, Kaupinis A, Valius M, Arlauskiene A, Navakauskiene R. Human amniotic fluid mesenchymal stem cells from second-and third-trimester amniocentesis: differentiation potential, molecular signature, and proteome analysis. Stem Cells Int. 2015; 2015:319238. DOI: 10.1155/2015/319238. PMID: 26351462. PMCID: PMC4553339.3. Guan X, Delo DM, Atala A, Soker S. In vitro cardiomyogenic potential of human amniotic fluid stem cells. J Tissue Eng Regen Med. 2011; 5:220–228. DOI: 10.1002/term.308. PMID: 20687122. PMCID: PMC2975013.
Article4. Gasiūnienė M, Zubova A, Utkus A, Navakauskienė R. Epigenetic and metabolic alterations in human amniotic fluid stem cells induced to cardiomyogenic differentiation by DNA methyltransferases and p53 inhibitors. J Cell Biochem. 2018; [Epub ahead of print]. DOI: 10.1002/jcb.28092. PMID: 30485506.
Article5. Maioli M, Contini G, Santaniello S, Bandiera P, Pigliaru G, Sanna R, Rinaldi S, Delitala AP, Montella A, Bagella L, Ventura C. Amniotic fluid stem cells morph into a cardiovascular lineage: analysis of a chemically induced cardiac and vascular commitment. Drug Des Devel Ther. 2013; 7:1063–1073. PMID: 24101862. PMCID: 3790833.6. Ahmadian E, Jafari S, Yari Khosroushahi A. Role of angiotensin II in stem cell therapy of cardiac disease. J Renin Angiotensin Aldosterone Syst. 2015; 16:702–711. DOI: 10.1177/1470320315621225. PMID: 26670032.
Article7. Xing Y, Lv A, Wang L, Yan X. The combination of angiotensin II and 5-azacytidine promotes cardiomyocyte differentiation of rat bone marrow mesenchymal stem cells. Mol Cell Biochem. 2012; 360:279–287. DOI: 10.1007/s11010-011-1067-z. PMID: 21935612.
Article8. Ikhapoh IA, Pelham CJ, Agrawal DK. Synergistic effect of angiotensin II on vascular endothelial growth factor-A-mediated differentiation of bone marrow-derived mesenchymal stem cells into endothelial cells. Stem Cell Res Ther. 2015; 6:4. DOI: 10.1186/scrt538. PMID: 25563650. PMCID: 4417220.
Article9. Azhar M, Schultz Jel J, Grupp I, Dorn GW 2nd, Meneton P, Molin DG, Gittenberger-de Groot AC, Doetschman T. Transforming growth factor beta in cardiovascular development and function. Cytokine Growth Factor Rev. 2003; 14:391–407. DOI: 10.1016/S1359-6101(03)00044-3. PMID: 12948523. PMCID: 3855389.
Article10. Mohanty S, Bose S, Jain KG, Bhargava B, Airan B. TGFβ1 contributes to cardiomyogenic-like differentiation of human bone marrow mesenchymal stem cells. Int J Cardiol. 2013; 163:93–99. DOI: 10.1016/j.ijcard.2011.08.003. PMID: 21903280.
Article11. Abdel-Latif A, Zuba-Surma EK, Case J, Tiwari S, Hunt G, Ranjan S, Vincent RJ, Srour EF, Bolli R, Dawn B. TGF-beta1 enhances cardiomyogenic differentiation of skeletal muscle-derived adult primitive cells. Basic Res Cardiol. 2008; 103:514–524. DOI: 10.1007/s00395-008-0729-9. PMID: 18500484. PMCID: 4270753.
Article12. Lv Y, Liu B, Wang HP, Zhang L. Intramyocardial implantation of differentiated rat bone marrow mesenchymal stem cells enhanced by TGF-β1 improves cardiac function in heart failure rats. Braz J Med Biol Res. 2016; 49:e5273. DOI: 10.1590/1414-431x20165273. PMID: 27254663. PMCID: PMC4932821.
Article13. Wenzel S, Henning K, Habbig A, Forst S, Schreckenberg R, Heger J, Maxeiner H, Schlüter KD. TGF-beta1 improves cardiac performance via up-regulation of laminin receptor 37/67 in adult ventricular cardiomyocytes. Basic Res Cardiol. 2010; 105:621–629. DOI: 10.1007/s00395-010-0108-1. PMID: 20556404.
Article14. Glemžaitė M, Navakauskienė R. Osteogenic differentiation of human amniotic fluid mesenchymal stem cells is determined by epigenetic changes. Stem Cells Int. 2016; 2016:6465307. DOI: 10.1155/2016/6465307. PMID: 27818691. PMCID: PMC5080506.
Article15. Johansson B. Low level operations and learning in computer vision [PhD dissertation]. Linköping: Linköping University;2004.16. Forrester SJ, Booz GW, Sigmund CD, Coffman TM, Kawai T, Rizzo V, Scalia R, Eguchi S. Angiotensin II signal transduction: an update on mechanisms of physiology and pathophysiology. Physiol Rev. 2018; 98:1627–1738. DOI: 10.1152/physrev.00038.2017. PMID: 29873596. PMCID: 6335102.
Article17. Wang MK, Sun HQ, Xiang YC, Jiang F, Su YP, Zou ZM. Different roles of TGF-β in the multi-lineage differentiation of stem cells. World J Stem Cells. 2012; 4:28–34. DOI: 10.4252/wjsc.v4.i5.28. PMID: 22993659. PMCID: 3443709.
Article18. Gaafar T, Shawky S, Attia W, Hamza H, El Hawary R. The role of angiotensin II in cardiomyogenic differentiation of human adipose tissue-derived mesenchymal stem cells. Comp Clin Pathol. 2015; 24:879–885. DOI: 10.1007/s00580-014-2001-z.
Article19. Liu BW, Lü AL, Hou J, Huang W, Hou H, Hou ZL, DAJ , Ai SY. Electrophysiological characteristics of cardiomyocyte-like cells from rat bone marrow derived mesenchymal stem cells by four inductors. Chin Med J (Engl). 2013; 126:3528–3533. PMID: 24034103.20. Li TS, Hayashi M, Ito H, Furutani A, Murata T, Matsuzaki M, Hamano K. Regeneration of infarcted myocardium by intramyocardial implantation of ex vivo transforming growth factor-beta-preprogrammed bone marrow stem cells. Circulation. 2005; 111:2438–2445. DOI: 10.1161/01.CIR.0000167553.49133.81. PMID: 15883211.
Article21. Honda M, Kiyokawa J, Tabo M, Inoue T. Electrophysiological characterization of cardiomyocytes derived from human induced pluripotent stem cells. J Pharmacol Sci. 2011; 117:149–159. DOI: 10.1254/jphs.11038FP. PMID: 22027094.
Article22. Cao L, Chen Y, Lu L, Liu Y, Wang Y, Fan J, Yin Y. Angiotensin II upregulates fibroblast-myofibroblast transition through Cx43-dependent CaMKII and TGF-β1 signaling in neonatal rat cardiac fibroblasts. Acta Biochim Biophys Sin. 2018; 50:843–852. DOI: 10.1093/abbs/gmy090. PMID: 30060053.
Article23. Boengler K, Schulz R, Heusch G. Connexin 43 signalling and cardioprotection. Heart. 2006; 92:1724–1727. DOI: 10.1136/hrt.2005.066878. PMID: 16387816. PMCID: 1861299.
Article24. Armiñán A, Gandía C, Bartual M, García-Verdugo JM, Lledó E, Mirabet V, Llop M, Barea J, Montero JA, Sepúlveda P. Cardiac differentiation is driven by NKX2.5 and GATA4 nuclear translocation in tissue-specific mesenchymal stem cells. Stem Cells Dev. 2009; 18:907–918. DOI: 10.1089/scd.2008.0292. PMID: 18983250.
Article25. Capasso S, Alessio N, Squillaro T, Di Bernardo G, Melone MA, Cipollaro M, Peluso G, Galderisi U. Changes in autophagy, proteasome activity and metabolism to determine a specific signature for acute and chronic senescent mesenchymal stromal cells. Oncotarget. 2015; 6:39457–39468. DOI: 10.18632/oncotarget.6277. PMID: 26540573. PMCID: 4741838.
Article26. Folmes CD, Dzeja PP, Nelson TJ, Terzic A. Metabolic plasticity in stem cell homeostasis and differentiation. Cell Stem Cell. 2012; 11:596–606. DOI: 10.1016/j.stem.2012.10.002. PMID: 23122287. PMCID: 3593051.
Article27. Rana P, Anson B, Engle S, Will Y. Characterization of human-induced pluripotent stem cell-derived cardiomyocytes: bioenergetics and utilization in safety screening. Toxicol Sci. 2012; 130:117–131. DOI: 10.1093/toxsci/kfs233. PMID: 22843568.
Article28. Denning C, Borgdorff V, Crutchley J, Firth KS, George V, Kalra S, Kondrashov A, Hoang MD, Mosqueira D, Patel A, Prodanov L, Rajamohan D, Skarnes WC, Smith JG, Young LE. Cardiomyocytes from human pluripotent stem cells: from laboratory curiosity to industrial biomedical platform. Biochim Biophys Acta. 2016; 1863:1728–1748. DOI: 10.1016/j.bbamcr.2015.10.014. PMID: 26524115. PMCID: PMC5221745.
Article29. Ventura-Clapier R, Garnier A, Veksler V. Transcriptional control of mitochondrial biogenesis: the central role of PGC-1alpha. Cardiovasc Res. 2008; 79:208–217. DOI: 10.1093/cvr/cvn098. PMID: 18430751.
Article30. Wu Z, Puigserver P, Andersson U, Zhang C, Adelmant G, Mootha V, Troy A, Cinti S, Lowell B, Scarpulla RC, Spiegelman BM. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell. 1999; 98:115–124. DOI: 10.1016/S0092-8674(00)80611-X. PMID: 10412986.
Article31. Xia Y, Buja LM, Scarpulla RC, McMillin JB. Electrical stimulation of neonatal cardiomyocytes results in the sequential activation of nuclear genes governing mitochondrial proliferation and differentiation. Proc Natl Acad Sci U S A. 1997; 94:11399–11404. DOI: 10.1073/pnas.94.21.11399. PMID: 9326621. PMCID: 23479.
Article32. Ng KM, Lee YK, Chan YC, Lai WH, Fung ML, Li RA, Siu CW, Tse HF. Exogenous expression of HIF-1 alpha promotes cardiac differentiation of embryonic stem cells. J Mol Cell Cardiol. 2010; 48:1129–1137. DOI: 10.1016/j.yjmcc.2010.01.015. PMID: 20116384.
Article33. Goumans MJ, de Boer TP, Smits AM, van Laake LW, van Vliet P, Metz CH, Korfage TH, Kats KP, Hochstenbach R, Pasterkamp G, Verhaar MC, van der Heyden MA, de Kleijn D, Mummery CL, van Veen TA, Sluijter JP, Doevendans PA. TGF-beta1 induces efficient differentiation of human cardiomyocyte progenitor cells into functional cardiomyocytes in vitro. Stem Cell Res. 2007; 1:138–149. DOI: 10.1016/j.scr.2008.02.003. PMID: 19383394.
Article34. De Coppi P, Bartsch G Jr, Siddiqui MM, Xu T, Santos CC, Perin L, Mostoslavsky G, Serre AC, Snyder EY, Yoo JJ, Furth ME, Soker S, Atala A. Isolation of amniotic stem cell lines with potential for therapy. Nat Biotechnol. 2007; 25:100–106. DOI: 10.1038/nbt1274. PMID: 17206138.
Article35. Klose RJ, Bird AP. Genomic DNA methylation: the mark and its mediators. Trends Biochem Sci. 2006; 31:89–97. DOI: 10.1016/j.tibs.2005.12.008. PMID: 16403636.
Article36. Liu Z, Li T, Liu Y, Jia Z, Li Y, Zhang C, Chen P, Ma K, Affara N, Zhou C. WNT signaling promotes Nkx2.5 expression and early cardiomyogenesis via downregulation of Hdac1. Biochim Biophys Acta. 2009; 1793:300–311. DOI: 10.1016/j.bbamcr.2008.08.013. PMID: 18851995.
Article37. Montgomery RL, Davis CA, Potthoff MJ, Haberland M, Fielitz J, Qi X, Hill JA, Richardson JA, Olson EN. Histone deacetylases 1 and 2 redundantly regulate cardiac morphogenesis, growth, and contractility. Genes Dev. 2007; 21:1790–1802. DOI: 10.1101/gad.1563807. PMID: 17639084. PMCID: 1920173.
Article38. Coppola A, Romito A, Borel C, Gehrig C, Gagnebin M, Falconnet E, Izzo A, Altucci L, Banfi S, Antonarakis SE, Minchiotti G, Cobellis G. Cardiomyogenesis is controlled by the miR-99a/let-7c cluster and epigenetic modifications. Stem Cell Res. 2014; 12:323–337. DOI: 10.1016/j.scr.2013.11.008. PMID: 24365598.
Article39. Zhou Y, Wang L, Vaseghi HR, Liu Z, Lu R, Alimohamadi S, Yin C, Fu JD, Wang GG, Liu J, Qian L. Bmi1 is a key epigenetic barrier to direct cardiac reprogramming. Cell Stem Cell. 2016; 18:382–395. DOI: 10.1016/j.stem.2016.02.003. PMID: 26942853. PMCID: 4779178.
Article40. Wang M, Yu Q, Wang L, Gu H. Distinct patterns of histone modifications at cardiac-specific gene promoters between cardiac stem cells and mesenchymal stem cells. Am J Physiol Cell Physiol. 2013; 304:C1080–C1090. DOI: 10.1152/ajpcell.00359.2012. PMID: 23552285.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Effects of TGF-β1 Overexpression on Biological Characteristics of Human Dental Pulp-derived Mesenchymal Stromal Cells
- Comparison of the Cardiomyogenic Potency of Human Amniotic Fluid and Bone Marrow Mesenchymal Stem Cells
- A Mini Overview of Isolation, Characterization and Application of Amniotic Fluid Stem Cells
- Cardiomyogenic Potential of Human Adipose Tissue and Umbilical Cord Derived-Mesenchymal Like Stem Cells
- Comparison with human amniotic membrane- and adipose tissue-derived mesenchymal stem cells
