Upregulation of PITX2 Promotes Letrozole Resistance Via Transcriptional Activation of IFITM1 Signaling in Breast Cancer Cells
- Affiliations
-
- 1Department of Breast Surgery, First Affiliated Hospital of China Medical University, Shenyang, China.
- 2Department of Medical Imaging, Cancer Hospital of China Medical University, Liaoning Cancer Hospital & Institute, Shenyang, China.
- 3Department of Breast Surgery, Cancer Hospital of China Medical University, Liaoning Cancer Hospital & Institute, Shenyang, China. zhangqiang8220@163.com
- KMID: 2464405
- DOI: http://doi.org/10.4143/crt.2018.100
Abstract
- PURPOSE
Although the interferon α (IFNα) signaling and the paired-like homeodomain transcription factor 2 (PITX2) have both been implicated in the progression of breast cancer (BCa), it remains obscure whether these two pathways act in a coordinated manner. We therefore aimed to elucidate the expression and function of PITX2 during the pathogenesis of endocrine resistance in BCa.
MATERIALS AND METHODS
PITX2 expression was assessed in BCa tissues using quantitative reverse transcription polymerase chain reaction (RT-qPCR) and immunohistochemistry and in experimentally induced letrozole-resistant BCa cells using RT-qPCR and immunoblotting. Effects of PITX2 deregulation on BCa progression was determined by assessing MTT, apoptosis and xenograft model. Finally, using multiple assays, the transcriptional regulation of interferon-inducible transmembrane protein 1 (IFITM1) by PITX2 was studied at both molecular and functional levels.
RESULTS
PITX2 expression was induced in letrozole-resistant BCa tissues and cells, and PITX2 induction by IFNα signaling powerfully protected BCa cells against letrozole insult and potentiated letrozole-resistance. Mechanistically, PITX2 enhanced IFNα-induced AKT activation by transactivating the transcription of IFITM1, thus rendering BCa cells unresponsive to letrozoleelicited cell death. Additionally, ablation of IFITM1 expression using siRNA substantially abolished IFNα-elicited AKT phosphorylation, even in the presence of PITX2 overexpression, thus sensitizing BCa cells to letrozole treatment.
CONCLUSION
These results demonstrate that constitutive upregulation of PITX2/IFITM1 cascade is an intrinsic adaptive mechanism during the pathogenesis of letrozole-resistance, and modulation of PITX2/IFITM1 level using different genetic and pharmacological means would thus have a novel therapeutic potential against letrozole resistance in BCa.
Keyword
MeSH Terms
Figure
-
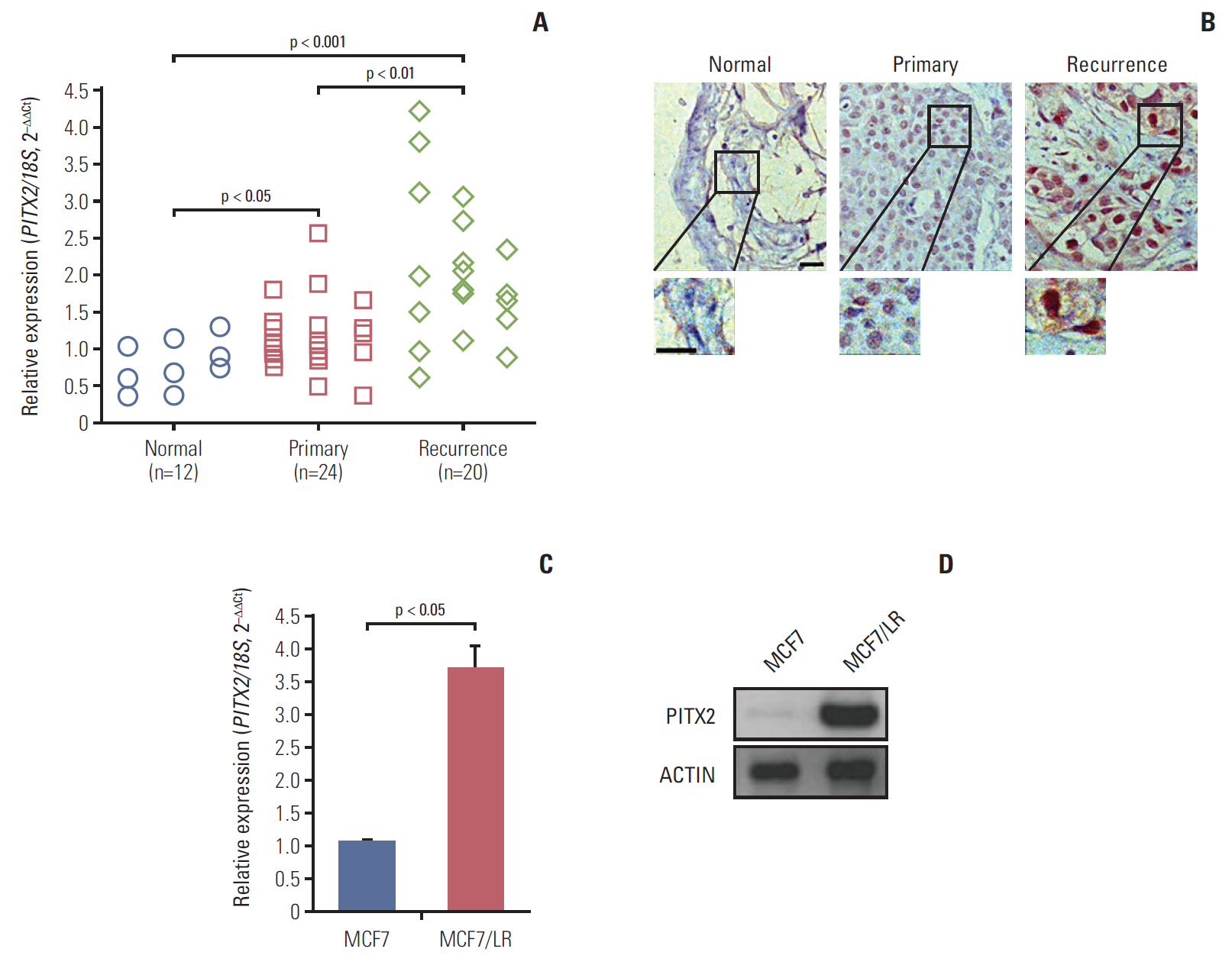
Fig. 1. Upregulation of paired-like homeodomain transcription factor 2 (PITX2) expression in letrozole-resistant breast cancer (BCa) tissues and cells. (A) PITX2 expression in normal breast tissues, primary BCa tissues and recurrent BCa tissues was determined using quantitative real-time polymerase chain reaction (RT-qPCR). Fold change was determined for each sample relative to the internal control gene 18S. Each value is a mean±standard error of mean from three experiments. (B) Representative fields of normal breast tissues, primary BCa tissues and recurrent BCa tissues immunohistochemically stained with anti-PITX2 antibody as indicated. Scale bar=20 μm. (C) Characterization of PITX2 expression in different BCa cells using RT-qPCR. (D) Western blotting analysis of PITX2 expression in different BCa cells. Actin served as the loading control.
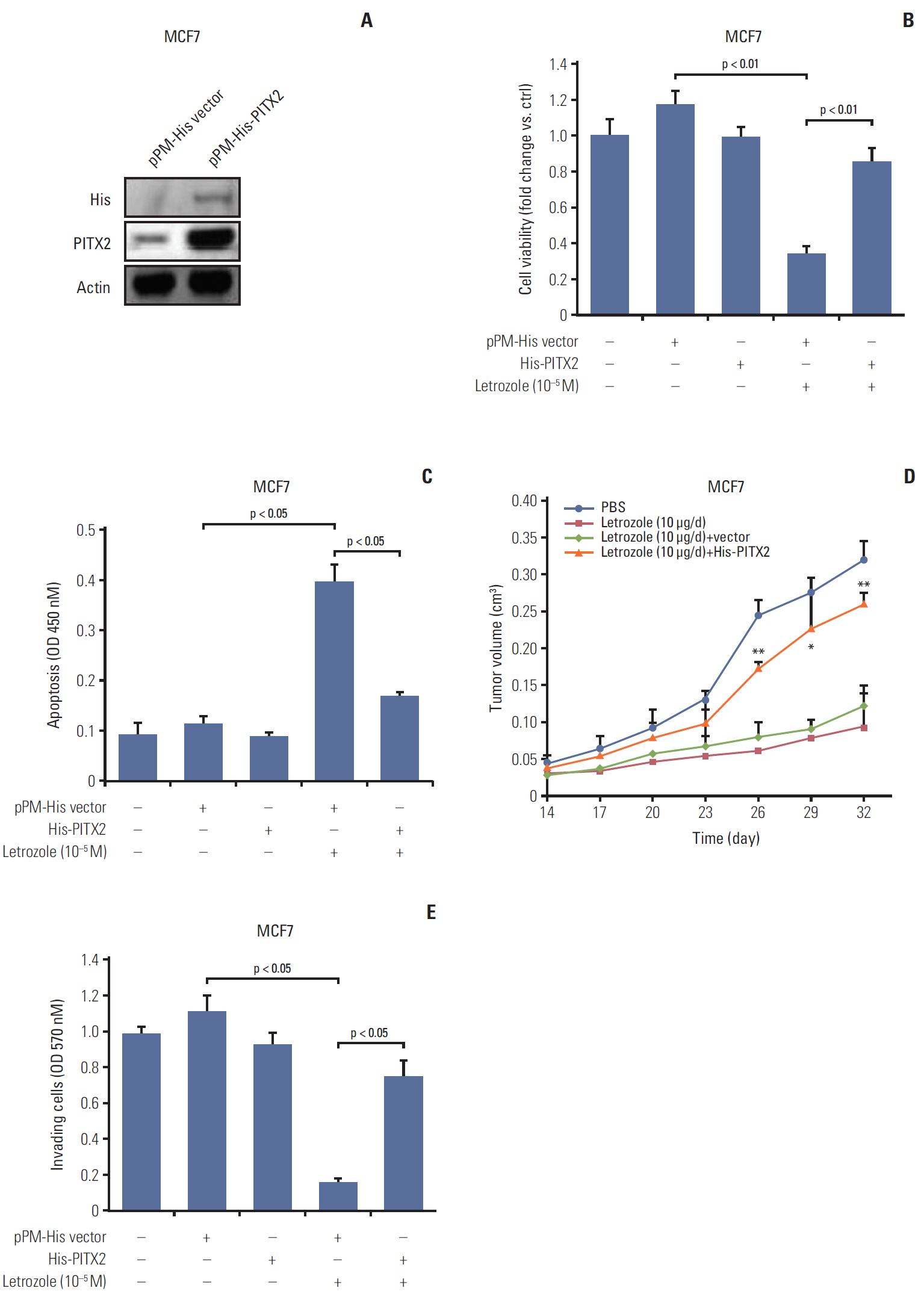
Fig. 2. Ectopic expression of exogenous paired-like homeodomain transcription factor 2 (PITX2) promotes letrozole resistance and cell invasiveness. (A) Western blotting analysis of PITX2 level in MCF7 cells stably transfected with pPM-His-PITX2 or pPM-His vector. After a 3-day culture and a following overnight starvation, MCF7 cells stably transfected with pPM-His-PITX2 or pPM-His vector were treated with 10–5 M of letrozole, along with 25 nM of androstenedione, for another 4 days. Viable cell numbers were then determined using trypan blue staining (B) (fold change was determined for each treatment relative to the untreated control cells, *p < 0.05 and **p < 0.01) and cell apoptosis was determined using an apoptosis enzyme-linked immunosorbent assay kit (C). The results were presented as the mean±standard error of mean of the triplicate samples. (D) Each mouse received subcutaneous injections at one site on each flank with 0.1 mL of suspension of different transfected MCF7 cells (2×107 cells/mL). Mice were then injected subcutaneously daily for 32 days with androstenedione (100 μg/day) plus letrozole (10 μg/day) from the day of inoculation. PBS, phosphate buffered saline. Tumor volumes were measured every 3 days. *p < 0.05 and **p < 0.01 when comparing letrozole (10 μg/day)+vector group to letrozole (10 μg/day)+His-PITX2 group. (E) Cell invasiveness assay: breast cancer cells (1×104) were cultured in the upper chamber of Transwell with a membrane coated with Matrigel for 24 hours, followed by treatment with 10–5 M of letrozole, along with 25 nM of Δ4A, or with dimethyl sulfoxide (Sigma-Aldrich) for 48 hours. The remaining cells on the membrane were then stained for 10 minutes with 0.1% crystal violet solution and subsequent spectrophotometry was developed at 570 nM.
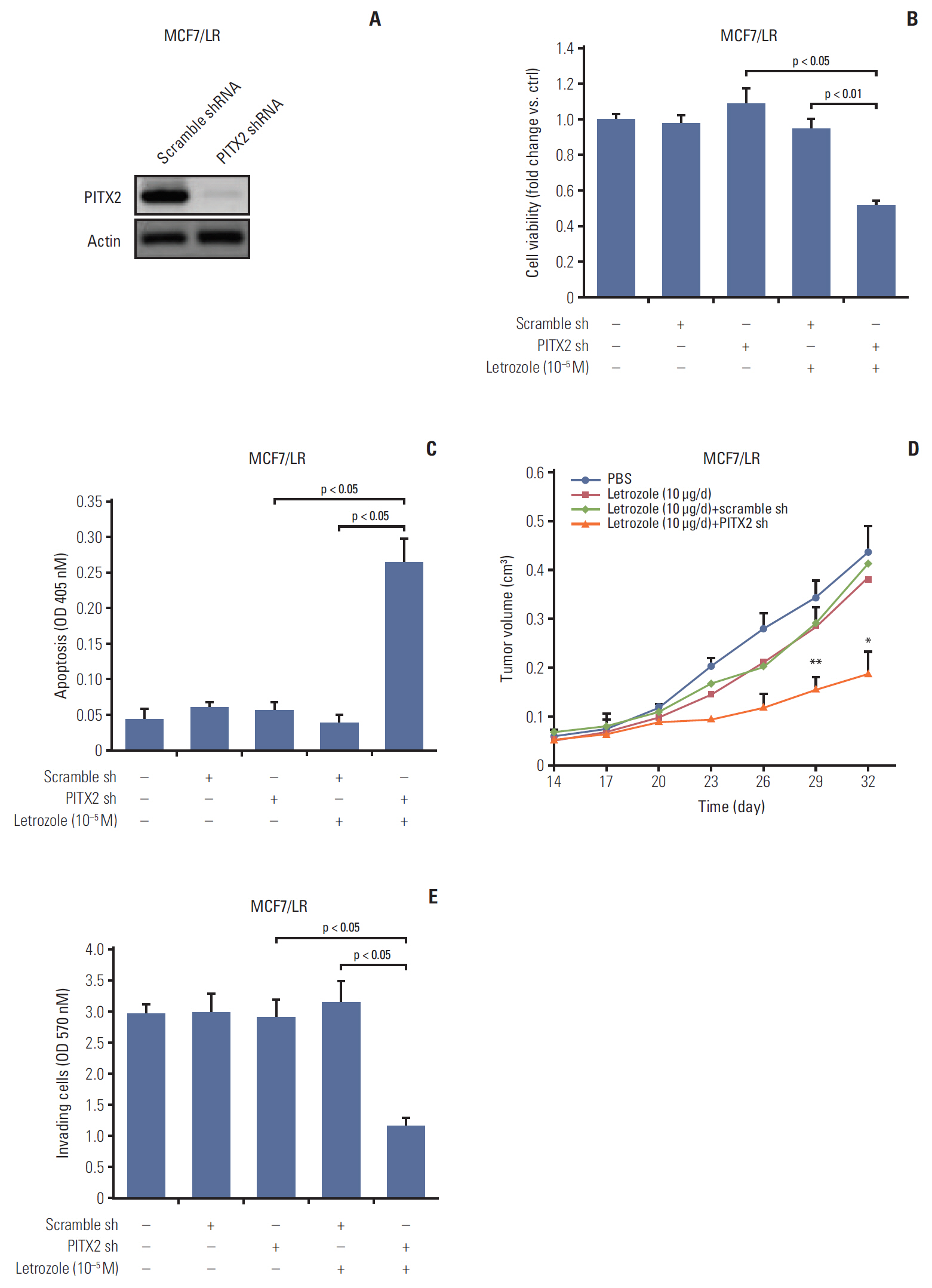
Fig. 3. Paired-like homeodomain transcription factor 2 (PITX2) inhibition sensitizes MCF7/LR cells to letrozole treatment. (A) MCF7/LR cells were transfected with PITX2 shRNA or scramble shRNA, followed by puromycin selection for 1-2 weeks. Stable knockdown of PITX2 expression was then verified by Western blotting analysis. Actin served as the loading control. (B) After a 3-day culture and a following overnight starvation, MCF7/LR cells stably transfected with PITX2 shRNA or scramble shRNA were treated with 10–5 M of letrozole, along with 25 nM of androstenedione, for another 4 days. Viable cell numbers were then determined using trypan blue staining. Fold change was determined for each treatment relative to the untreated control cells. (C) After a 3-day culture and a following overnight starvation, MCF7 cells stably transfected with pPM-His-PITX2 or pPM-His vector were treated with 10–5 M of letrozole, along with 25 nM of androstenedione, for another 4 days. Cell apoptosis was determined using an apoptosis enzyme-linked immunosorbent assay kit. (D) Tumor xenograft assay, as described above, was carried out to evaluate the effects of PITX2 inhibition on in vivo letrozole sensitivity. *p < 0.05 and **p < 0.01 when comparing letrozole (10 μg/day)+scramble shRNA to letrozole (10 μg/day)+PITX2 shRNA. (E) Cell invasiveness assay as described above. Shown are the absorptions at 570 nm of MCF7/LR cells treated with different transfection plus 10–5 M of letrozole for 48 hours, in the lower chambers of transwells.
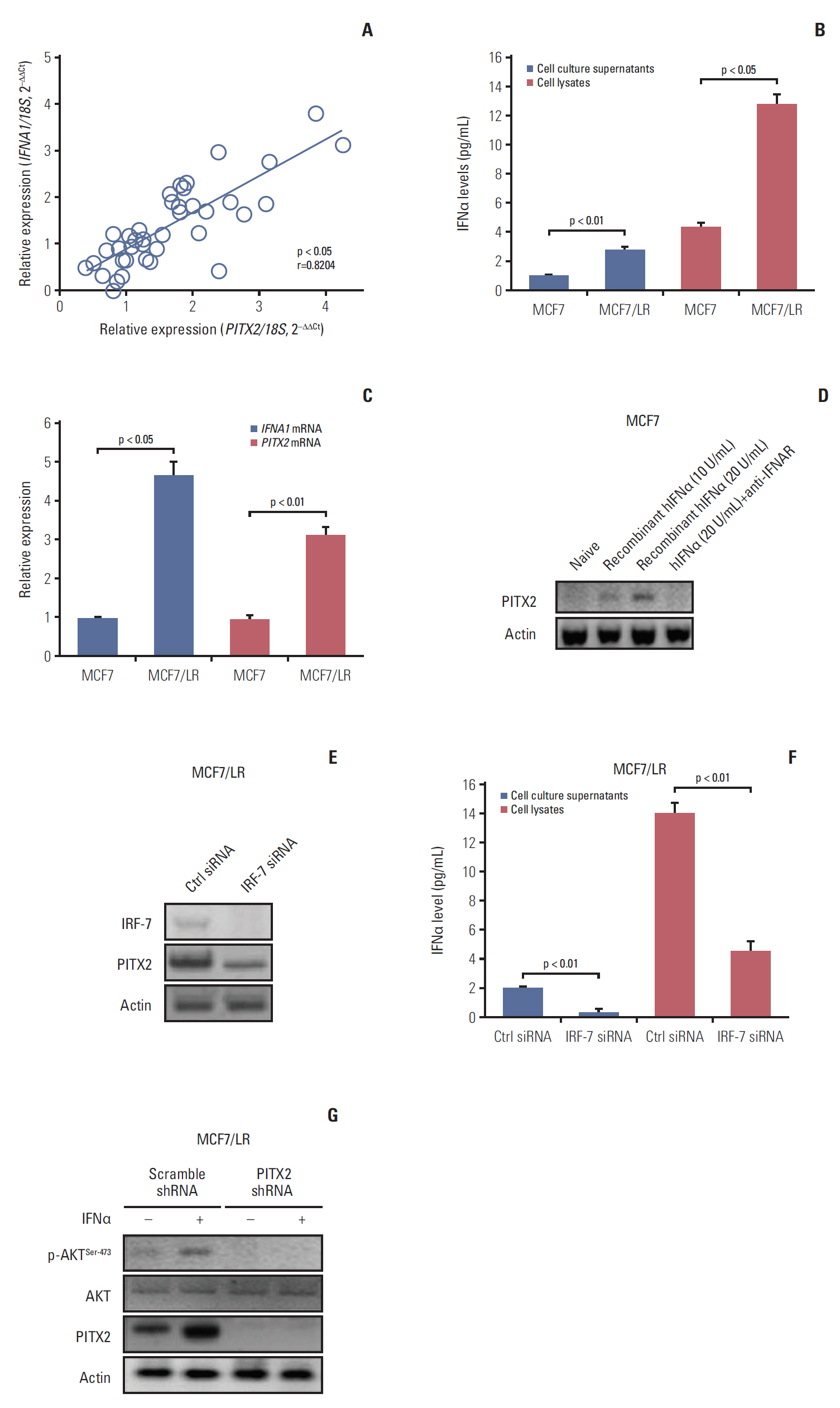
Fig. 4. Activation of interferon α (IFNα) signaling pathway stimulates paired-like homeodomain transcription factor 2 (PITX2) expression in letrozole-resistant breast cancer (BCa) cells. (A) PITX2 and IFNA1 expression levels in primary BCa tissues (n=24) and recurrent BCa tissues (n=20) were determined using quantitative real-time polymerase chain reaction (RT-qPCR). Fold change was determined for each sample relative to the internal control gene 18S. Each value is a mean±standard error of mean from three experiments. The correlation between PITX2 and IFNA1 expression levels were the analyzed using Pearson chi-square test. (B) Enzyme-linked immunosorbent assay (ELISA) analysis of baseline expression of IFNα in cell lysates and culture supernatants in different BCa cells. (C) Relative expression levels of PITX2 and IFNA1 mRNA were assayed using RT-qPCR in different BCa cells. (D) MCF7 cells were challenged for 24 hours with a gradual concentration of recombinant hIFNα protein, in the presence or absence of the pretreatment with 5 μg/mL of anti-IFNAR neutralizing antibody for 4 hours. PITX2 expression was then determined using Western blotting. Actin served as the loading control. (E) MCF7/LR cells were transiently transfected with IRF-7 siRNA or Ctrl siRNA using Lipofectamine 3000. Forty-eight hours later, cells were harvested and subjected to Western blotting analysis of IRF-7 and PITX2 expression. (F) ELISA analysis of baseline expression of IFNα in cell lysates and culture supernatants in MCF7/LR cells with different transfections. (G) MCF7/LR cells with different transfections were challenged with 20 U/mL of recombinant hIFNα protein for 24 hours, followed by Western blotting analysis of p-AKTSer-473, AKT and PITX2 expression.
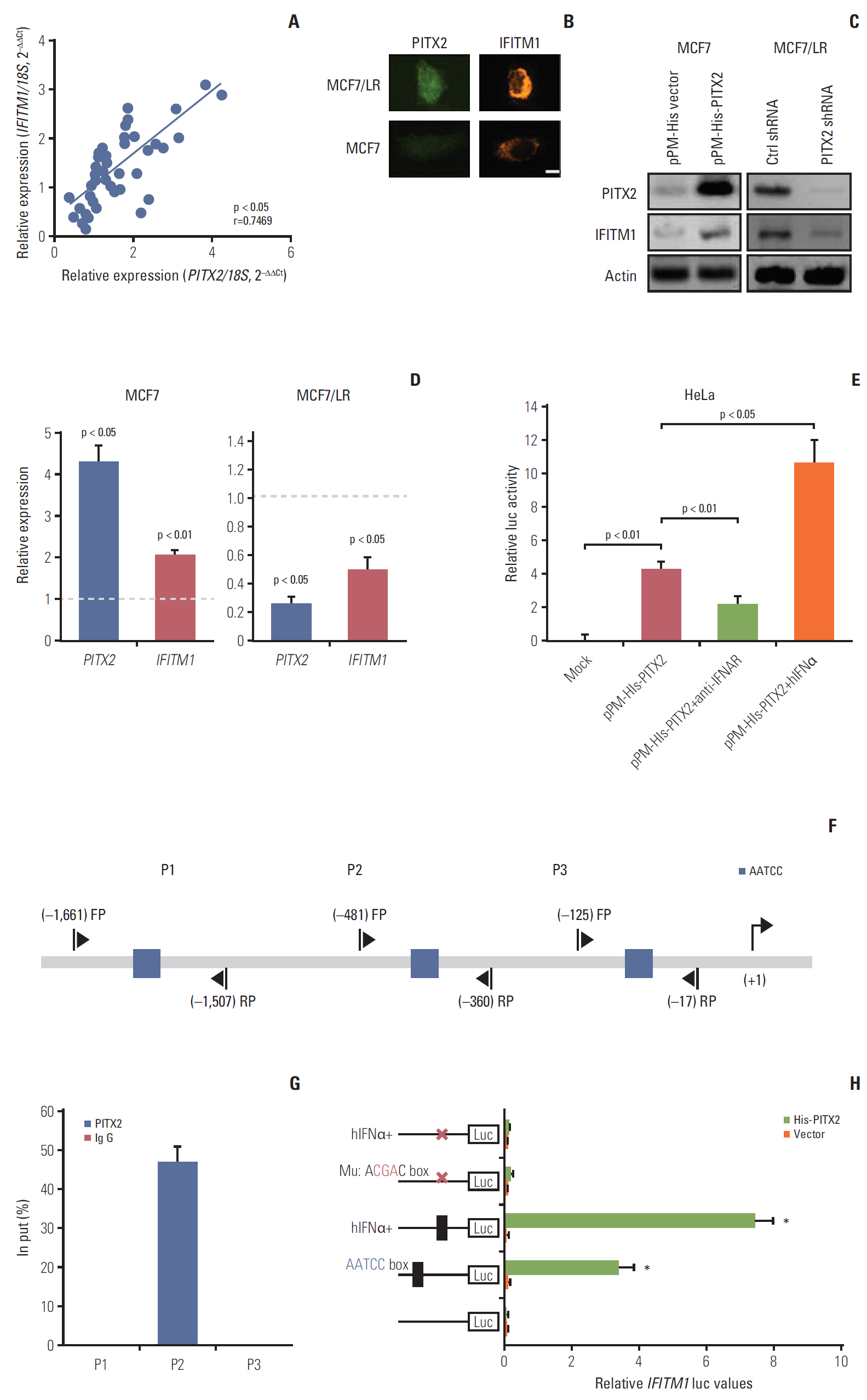
Fig. 5. Direct regulation of interferon-inducible transmembrane protein 1 (IFITM1) transcription by paired-like homeodomain transcription factor 2 (PITX2) in breast cancer (BCa) cells. (A) PITX2 and IFITM1 expression levels in primary BCa tissues (n=24) and recurrent BCa tissues (n=20) were determined using quantitative real-time polymerase chain reaction (RT-qPCR). Fold change was determined for each sample relative to the internal control gene 18S. Each value is a mean±standard error of mean from three experiments. The correlation between PITX2 and IFITM1 expression levels were the analyzed using Pearson chi-square test. (B) Iimmunofluorescence microscopy was carried out by co-staining MCF7 and MCF7/LR cells with anti-PITX2 (green) and anti-IFITM1 (red) antibodies. Scale bar=2.5 μm. (C) Western blotting analysis of PITX2 and IFITM1 levels in MCF7/His-PITX2, MCF7/His-vector, MCF7/LR/PITX2 shRNA and MCF7/LR/scramble shRNA cells. Actin served as the loading control. (D) RT-qPCR analysis of PITX2 and IFITM1 levels in MCF7/His-PITX2, MCF7/His-vector, MCF7/LR/PITX2 shRNA, and MCF7/LR/scramble shRNA cells. Fold change was determined for each gene relative to the values in Ctrl cells (dashed line). (E) The pGL4-Luc-IFITM1 reporter plasmid and pRL-TK Renilla reporter plasmid were co-transfected with pPM-His-PITX2 or pPM-His vector into HeLa cells using FUGENE. Forty-eight hours later, cells were treated for 4 hours with 20 U/mL of hIFNα or with 5 μg/mL of anti-IFNAR neutralizing antibody, followed by measurement of luciferase activity. (F) Simplified structure of the potential binding site of PITX2 onto IFITM1 promoter. (G) MCF7/His-PITX2 cells were challenged for 4 hours with 20 U/mL of hIFNα, followed by chromatin immunoprecipitation and quantitative polymerase chain reaction analysis of recruitment of PITX2 onto IFITM1 promoter. (H) The mutation of the putative IFITM1-binding site on the promoter at –406/–402 was carried out by replacing AATCC with AcgaC using the QuikChange Site-Directed Mutagenesis Kit. The wild type or mutated pGL4-Luc-IFITM1 reporter plasmid and pRL-TK Renilla reporter plasmid were co-transfected with pPM-His-PITX2 or pPM-His vector into HeLa cells using FUGENE. Forty-eight hours later, cells were treated for 4 hours with 20 U/mL of hIFNα. Relative luciferase activity was then measured using a dual luciferase reporter assay.
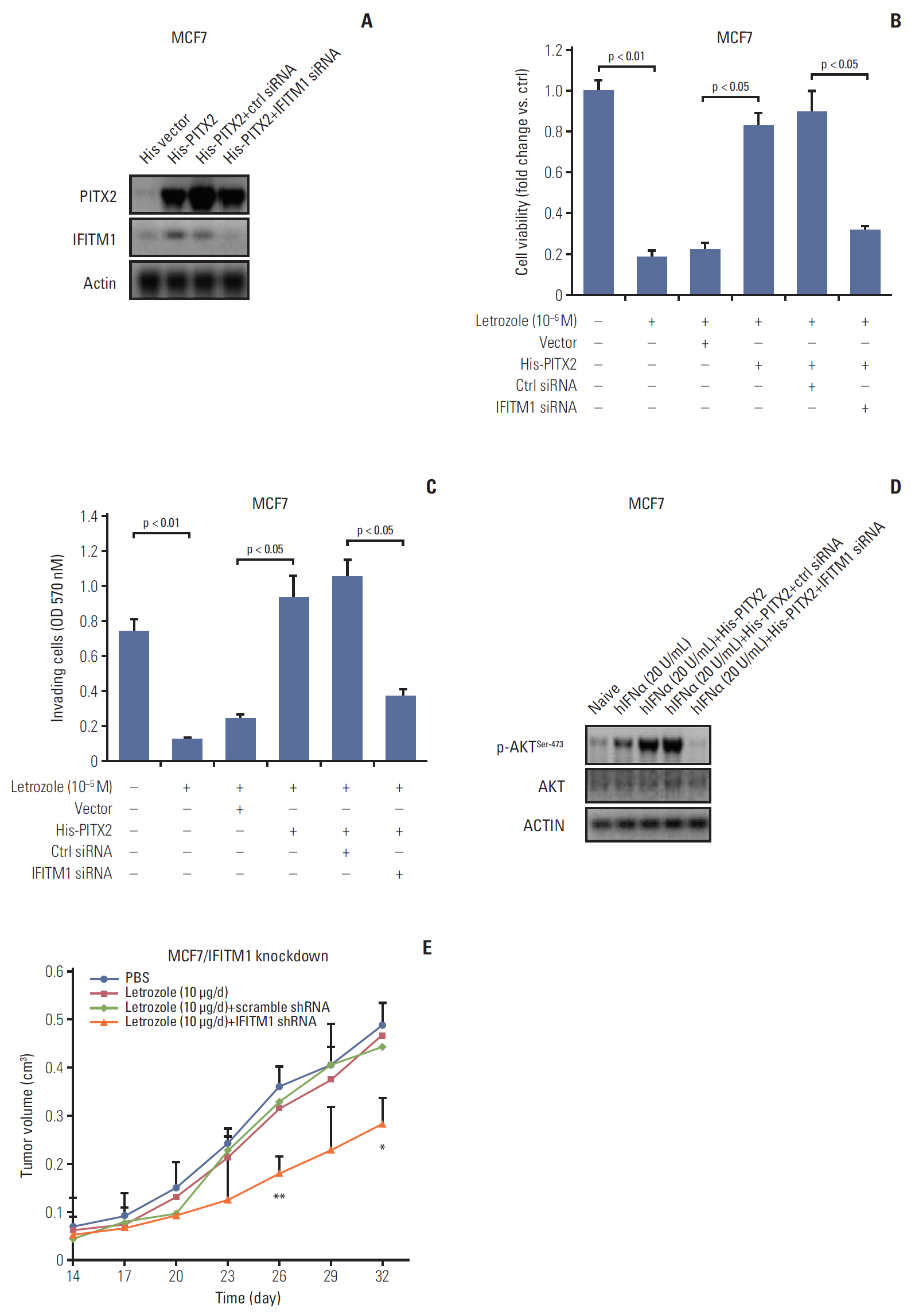
Fig. 6. Transient knockdown of interferon-inducible transmembrane protein 1 (IFITM1) abolishes paired-like homeodomain transcription factor 2 (PITX2)‒overexpression induced letrozole-resistance. (A) MCF7/LR/His-PITX2 cells were transiently transfected with IFITM1 siRNA or Ctrl siRNA. Forty-eight hours later, cells were harvested and subjected to Western blotting analysis of PITX2 and IFITM1 levels. After a 3-day culture and a following overnight starvation, the breast cancer (BCa) cells with different transfections were treated with 10–5 M of letrozole, along with 25 nM of androstenedione, for another 4 days. Viable cell numbers were then determined using trypan blue staining (B) (fold change was determined for each treatment relative to the untreated control cells) and cell invasiveness was determined using a colorimetric kit at 570 nm (C). The results were presented as the mean±standard error of mean of the triplicate samples. (D) BCa cells with different transfections were challenged with 20 U/mL of recombinant hIFNα protein for 24 hours, followed by Western blotting analysis of p-AKTSer-473 and AKT expression levels. (E) Tumor xenograft assay, as described above, was carried out to evaluate the effects of IFITM1 inhibition on in vivo letrozole sensitivity. *p < 0.05 and **p < 0.01 when comparing letrozole (10 μg/day)+scramble shRNA to letrozole (10 μg/day) +IFITM1 shRNA.
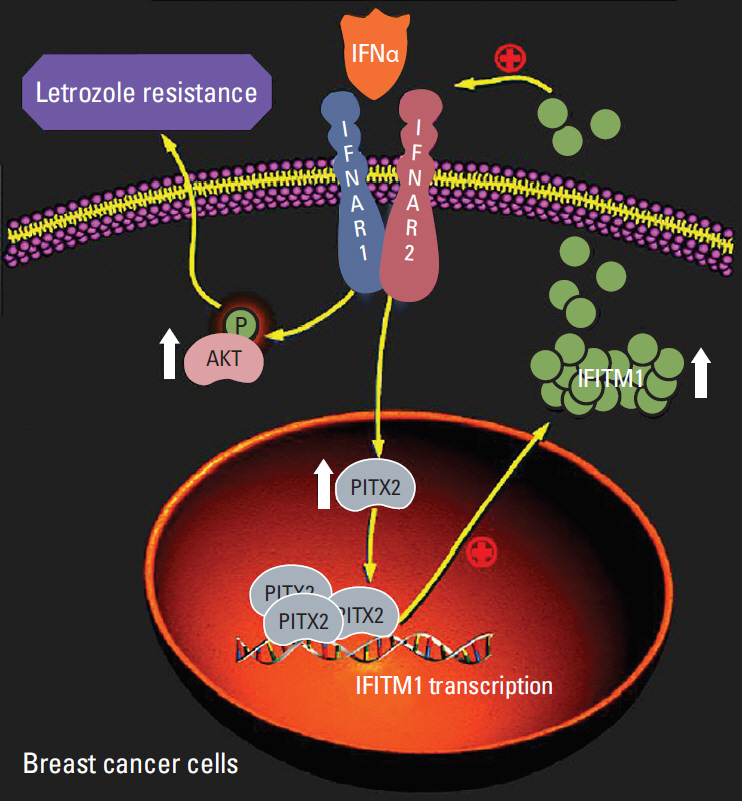
Fig. 7. Proposed working model in the current study.
Reference
-
References
1. Hoeflich KP, Guan J, Edgar KA, O'Brien C, Savage H, Wilson TR, et al. The PI3K inhibitor taselisib overcomes letrozole resistance in a breast cancer model expressing aromatase. Genes Cancer. 2016; 7:73–85.
Article2. Jelovac D, Sabnis G, Long BJ, Macedo L, Goloubeva OG, Brodie AM. Activation of mitogen-activated protein kinase in xenografts and cells during prolonged treatment with aromatase inhibitor letrozole. Cancer Res. 2005; 65:5380–9.
Article3. Brodie AM, Chumsri S, Sukumar S, Sabnis GJ. Extending aromatase inhibitor sensitivity in hormone resistant breast cancer. Horm Mol Biol Clin Investig. 2011; 5:97–103.
Article4. Choi HJ, Lui A, Ogony J, Jan R, Sims PJ, Lewis-Wambi J. Targeting interferon response genes sensitizes aromatase inhibitor resistant breast cancer cells to estrogen-induced cell death. Breast Cancer Res. 2015; 17:6.
Article5. Ariazi EA, Cunliffe HE, Lewis-Wambi JS, Slifker MJ, Willis AL, Ramos P, et al. Estrogen induces apoptosis in estrogen deprivation-resistant breast cancer through stress responses as identified by global gene expression across time. Proc Natl Acad Sci U S A. 2011; 108:18879–86.
Article6. Zacharias AL, Lewandoski M, Rudnicki MA, Gage PJ. Pitx2 is an upstream activator of extraocular myogenesis and survival. Dev Biol. 2011; 349:395–405.
Article7. Dietrich D, Hasinger O, Liebenberg V, Field JK, Kristiansen G, Soltermann A. DNA methylation of the homeobox genes PITX2 and SHOX2 predicts outcome in non-small-cell lung cancer patients. Diagn Mol Pathol. 2012; 21:93–104.
Article8. Lian ZQ, Wang Q, Li WP, Zhang AQ, Wu L. Screening of significantly hypermethylated genes in breast cancer using microarray-based methylated-CpG island recovery assay and identification of their expression levels. Int J Oncol. 2012; 41:629–38.
Article9. Vasiljevic N, Ahmad AS, Carter PD, Fisher G, Berney DM, Foster CS, et al. DNA methylation of PITX2 predicts poor survival in men with prostate cancer. Biomark Med. 2014; 8:1143–50.10. Zhang JX, Tong ZT, Yang L, Wang F, Chai HP, Zhang F, et al. PITX2: a promising predictive biomarker of patients' prognosis and chemoradioresistance in esophageal squamous cell carcinoma. Int J Cancer. 2013; 132:2567–77.
Article11. Lee WK, Chakraborty PK, Thevenod F. Pituitary homeobox 2 (PITX2) protects renal cancer cell lines against doxorubicin toxicity by transcriptional activation of the multidrug transporter ABCB1. Int J Cancer. 2013; 133:556–67.
Article12. Yang Z, Li C, Fan Z, Liu H, Zhang X, Cai Z, et al. Single-cell sequencing reveals variants in ARID1A, GPRC5A and MLL2 driving self-renewal of human bladder cancer stem cells. Eur Urol. 2017; 71:8–12.
Article13. Gu F, Hsu HK, Hsu PY, Wu J, Ma Y, Parvin J, et al. Inference of hierarchical regulatory network of estrogen-dependent breast cancer through ChIP-based data. BMC Syst Biol. 2010; 4:170.
Article14. Yu T, Yu HR, Sun JY, Zhao Z, Li S, Zhang XF, et al. miR-1271 inhibits ERalpha expression and confers letrozole resistance in breast cancer. Oncotarget. 2017; 8:107134–48.15. Dong YS, Hou WG, Li Y, Liu DB, Hao GZ, Zhang HF, et al. Unexpected requirement for a binding partner of the syntaxin family in phagocytosis by murine testicular Sertoli cells. Cell Death Differ. 2016; 23:787–800.
Article16. Zhang C, Lai JH, Hu B, Zhang S, Zhao J, Li W. A chromatin modifier regulates Sertoli cell response to mono-(2-ethylhexyl) phthalate (MEHP) via tissue inhibitor of metalloproteinase 2 (TIMP2) signaling. Biochim Biophys Acta. 2014; 1839:1170–82.
Article17. Zhang Q, Liu XY, Li S, Zhao Z, Li J, Cui MK, et al. Repression of ESR1 transcription by MYOD potentiates letrozole-resistance in ERalpha-positive breast cancer cells. Biochem Biophys Res Commun. 2017; 492:425–33.18. Ning P, Zhong JG, Jiang F, Zhang Y, Zhao J, Tian F, et al. Role of protein S in castration-resistant prostate cancer-like cells. Endocr Relat Cancer. 2016; 23:595–607.
Article19. Lopez JI, Angulo JC, Martin A, Sanchez-Chapado M, Gonzalez-Corpas A, Colas B, et al. A DNA hypermethylation profile reveals new potential biomarkers for the evaluation of prognosis in urothelial bladder cancer. APMIS. 2017; 125:787–96.
Article20. Kapoor S. Promising, new prognostic markers of esophageal carcinomas. APMIS. 2013; 121:1011.
Article21. Liu Y, Nan F, Lu K, Wang Y, Liu Y, Wei S, et al. Identification of key genes in endometrioid endometrial adenocarcinoma via TCGA database. Cancer Biomark. 2017; 21:11–21.
Article22. Vela I, Morrissey C, Zhang X, Chen S, Corey E, Strutton GM, et al. PITX2 and non-canonical Wnt pathway interaction in metastatic prostate cancer. Clin Exp Metastasis. 2014; 31:199–211.
Article23. Wan Abdul Rahman WF, Fauzi MH, Jaafar H. Expression of DNA methylation marker of paired-like homeodomain transcription factor 2 and growth receptors in invasive ductal carcinoma of the breast. Asian Pac J Cancer Prev. 2014; 15:8441–5.24. Jezkova E, Kajo K, Zubor P, Grendar M, Malicherova B, Mendelova A, et al. Methylation in promoter regions of PITX2 and RASSF1A genes in association with clinicopathological features in breast cancer patients. Tumour Biol. 2014; 37:15707–18.
Article25. Gultekin M, Eren G, Babacan T, Yildiz F, Altundag K, Guler N, et al. Metaplastic breast carcinoma: a heterogeneous disease. Asian Pac J Cancer Prev. 2014; 15:2851–6.
Article26. Khan MS, Pandith AA, Masoodi SR, Wani KA, Ul Hussain M, Mudassar S. Epigenetic silencing of TSHR gene in thyroid cancer patients in relation to their BRAF V600E mutation status. Endocrine. 2014; 47:449–55.
Article27. Zhang JX, Chen ZH, Xu Y, Chen JW, Weng HW, Yun M, et al. Downregulation of microRNA-644a promotes esophageal squamous cell carcinoma aggressiveness and stem cell-like phenotype via dysregulation of PITX2. Clin Cancer Res. 2017; 23:298–310.
Article28. Torrado M, Franco D, Lozano-Velasco E, Hernandez-Torres F, Calvino R, Aldama G, et al. A microRNA-transcription factor blueprint for early atrial arrhythmogenic remodeling. Biomed Res Int. 2015; 2015:263151.
Article29. Cavazzoni A, Bonelli MA, Fumarola C, La Monica S, Airoud K, Bertoni R, et al. Overcoming acquired resistance to letrozole by targeting the PI3K/AKT/mTOR pathway in breast cancer cell clones. Cancer Lett. 2012; 323:77–87.
Article30. Lei H, Furlong PJ, Ra JH, Mullins D, Cantor R, Fraker DL, et al. AKT activation and response to interferon-beta in human cancer cells. Cancer Biol Ther. 2005; 4:709–15.31. Li P, Shi ML, Shen WL, Zhang Z, Xie DJ, Zhang XY, et al. Coordinated regulation of IFITM1, 2 and 3 genes by an IFN-responsive enhancer through long-range chromatin interactions. Biochim Biophys Acta Gene Regul Mech. 2017; 1860:885–93.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- An Effect of Letrozole on Gastric Cancer?
- High Levels of Hyaluronic Acid Synthase-2 Mediate NRF2-Driven Chemoresistance in Breast Cancer Cells
- p90RSK Activation Promotes Epithelial-Mesenchymal Transition in Cisplatin-Treated Triple-Negative Breast Cancer Cells
- New Findings on Breast Cancer Stem Cells: A Review
- Nkx-2.5 Regulates MDR1 Expression via Its Upstream Promoter in Breast Cancer Cells
