Rhodanthpyrone A and B play an anti-inflammatory role by suppressing the nuclear factor-κB pathway in macrophages
- Affiliations
-
- 1College of Pharmacy, Woosuk University, Wanju 55338, Korea.
- 2Department of Pharmacology, School of Medicine, Wonkwang University, Iksan 54538, Korea.
- 3College of Pharmacy, Dankook University, Cheonan 31116, Korea.
- 4College of Pharmacy, Chonbuk National University, Jeonju 54896, Korea. ejbae7@jbnu.ac.kr
- KMID: 2461042
- DOI: http://doi.org/10.4196/kjpp.2019.23.6.493
Abstract
- Macrophage-associated inflammation is crucial for the pathogenesis of diverse diseases including metabolic disorders. Rhodanthpyrone (Rho) is an active component of Gentiana rhodantha, which has been used in traditional Chinese medicine to treat inflammation. Although synthesis procedures of RhoA and RhoB were reported, the biological effects of the specific compounds have never been explored. In this study, the anti-inflammatory activity and mechanisms of action of RhoA and RhoB were studied in lipopolysaccharide (LPS)-stimulated macrophages. Pretreatment with RhoA and RhoB decreased inducible nitric oxide synthase and cyclooxygenase-2 expressions in RAW 264.7 cells and in thioglycollate-elicited mouse peritoneal macrophages. In addition, it downregulated transcript levels of several inflammatory genes in LPS-stimulated RAW 264.7 cells, including inflammatory cytokines/chemokines (Tnfa, Il6, and Ccl2) and inflammatory mediators (Nos2 and Ptgs2). Macrophage chemotaxis was also inhibited by treatment with the compounds. Mechanistic studies revealed that RhoA and RhoB suppressed the nuclear factor (NF)-κB pathway, but not the canonical mitogen activated protein kinase pathway, in LPS-stimulated condition. Moreover, the inhibitory effect of RhoA and RhoB on inflammatory gene expressions was attenuated by treatment with an NF-κB inhibitor. Our findings suggest that RhoA and RhoB play an anti-inflammatory role at least in part by suppressing the NF-κB pathway during macrophage-mediated inflammation.
MeSH Terms
Figure
-

Fig. 1 Chemical structures of Rhodanthpyrone (Rho)A and RhoB, and the effects of RhoA and RhoB on the viability of RAW 264.7 cells. (A) Chemical structures of RhoA and RhoB. In (B), the results are expressed as the percentage of surviving cells treated with RhoA or RhoB for 24 h relative to the unstimulated surviving RAW 264.7 cells (n = 3). Values are expressed as mean ± SEM.

Fig. 2 Prevention of lipopolysaccharide (LPS)-stimulated inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) expression by Rhodanthpyrone (Rho)A and RhoB. (A) RAW 264.7 cells were pretreated with the indicated concentrations of RhoA or RhoB for 1 h, and then 10 ng/ml LPS was added for 24 h. Nitric oxide (NO) levels in the culture supernatant were determined (n = 3). (B) iNOS and COX-2 protein levels were analyzed in RAW 264.7 cells treated with LPS (10 ng/ml) in the presence or absence of RhoA or RhoB as indicated (n = 5). (C, D) RAW 264.7 cells were exposed to LPS (10 ng/ml) with or without RhoA or RhoB as indicated, and the protein (C) and mRNA (D) levels of iNOS and COX-2 were measured (n = 5). (E) Intraperitoneal macrophages were isolated as described in “Methods”. iNOS and COX-2 protein levels after stimulation with LPS (10 ng/ml) in the presence or absence of RhoA or RhoB were determined. Values are expressed as mean ± SEM. **p < 0.01 vs. untreated control; #p < 0.05 and ##p < 0.01 vs. LPS alone.
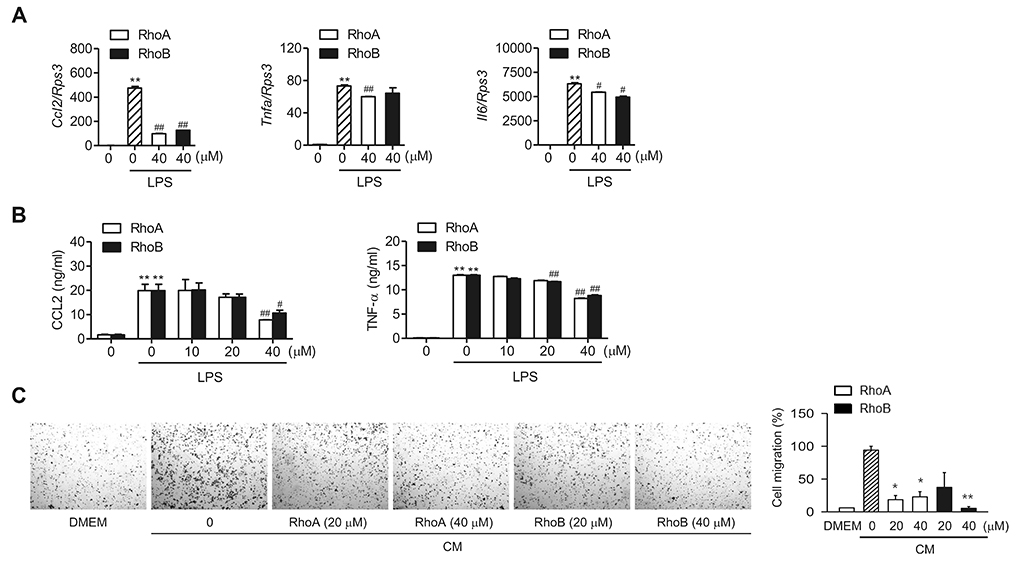
Fig. 3 Suppression of lipopolysaccharide (LPS)-stimulated pro-inflammatory cytokines/chemokine production by Rhodanthpyrone (Rho) A and RhoB. RAW 264.7 cells were treated with indicated concentrations of RhoA or RhoB for 1 h and then stimulated with 10 ng/ml LPS for 24 h. (A) mRNA levels of indicated genes were determined using qPCR (n = 3). (B) CCL2 and tumor necrosis factor (TNF)-α protein levels in the culture supernatant were determined using enzyme-linked immunosorbent assays (ELISA) (n = 3). (C) Representative images of the migration assay after staining cells with crystal violet (left). Cells that migrated to the bottom of the inserts were counted, and the quantification results are expressed as the relative ratio (%) (right). CM, conditioned medium. Values are expressed as mean ± SEM. *p < 0.05 and **p < 0.01 vs. LPS alone.
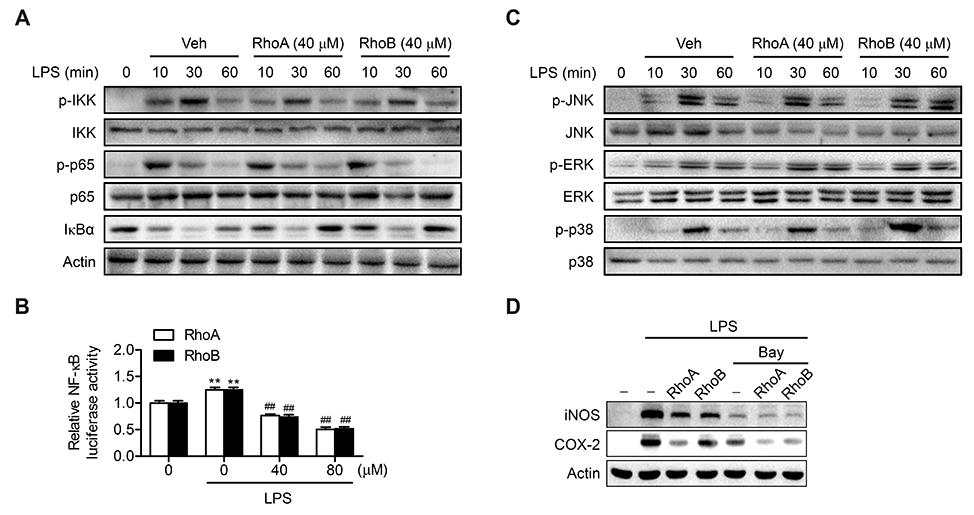
Fig. 4 Suppression of lipopolysaccharide (LPS)-stimulated nuclear factor (NF)-κB pathway by Rhodanthpyrone (Rho)A and RhoB. (A) RAW 264.7 cells were treated with 40 µM RhoA or RhoB for 1 h and then stimulated with 10 ng/ml LPS for 24 h. Phosphorylated- and total-inhibitory κB kinase (IKK), p65, and inhibitory κB kinase α (IκBα) were determined using western blotting. (B) RAW 264.7 cells transfected with NF-κB luciferase were treated with LPS alone, or LPS with RhoA or RhoB, and then NF-κB luciferase activity in cell lysates were assayed (n = 5–6). (C) The levels of phosphorylated-and total mitogen activated protein kinases (MAPKs) were analyzed in RAW 264.7 cells treated as in panel A. (D) RAW 264.7 cells were treated with RhoA or RhoB (40 µM, 1 h) in the presence or absence of Bay 11-7082 (5 mM), and then stimulated with LPS (10 ng/ml, 24 h). Values are expressed as mean ± SEM. **p < 0.01 vs. untreated control; ##p < 0.01 vs. LPS alone.
Reference
-
1. Okin D, Medzhitov R. Evolution of inflammatory diseases. Curr Biol. 2012; 22:R733–R740.
Article2. Wynn TA, Chawla A, Pollard JW. Macrophage biology in development, homeostasis and disease. Nature. 2013; 496:445–455.
Article3. Schultze JL, Schmieder A, Goerdt S. Macrophage activation in human diseases. Semin Immunol. 2015; 27:249–256.
Article4. de Jesus AA, Canna SW, Liu Y, Goldbach-Mansky R. Molecular mechanisms in genetically defined autoinflammatory diseases: disorders of amplified danger signaling. Annu Rev Immunol. 2015; 33:823–874.
Article5. Vallabhapurapu S, Karin M. Regulation and function of NF-kappaB transcription factors in the immune system. Annu Rev Immunol. 2009; 27:693–733.6. Beinke S, Ley SC. Functions of NF-kappaB1 and NF-kappaB2 in immune cell biology. Biochem J. 2004; 382(Pt 2):393–409.7. Xie QW, Kashiwabara Y, Nathan C. Role of transcription factor NF-kappa B/Rel in induction of nitric oxide synthase. J Biol Chem. 1994; 269:4705–4708.
Article8. Appleby SB, Ristimäki A, Neilson K, Narko K, Hla T. Structure of the human cyclo-oxygenase-2 gene. Biochem J. 1994; 302(Pt 3):723–727.
Article9. Guha M, Mackman N. LPS induction of gene expression in human monocytes. Cell Signal. 2001; 13:85–94.
Article10. Xu M, Zhang M, Wang D, Yang CR, Zhang YJ. Phenolic compounds from the whole plants of Gentiana rhodantha (Gentianaceae). Chem Biodivers. 2011; 8:1891–1900.
Article11. Oh DY, Talukdar S, Bae EJ, Imamura T, Morinaga H, Fan W, Li P, Lu WJ, Watkins SM, Olefsky JM. GPR120 is an omega-3 fatty acid receptor mediating potent anti-inflammatory and insulin-sensitizing effects. Cell. 2010; 142:687–698.
Article12. Ka SO, Song MY, Bae EJ, Park BH. Myeloid SIRT1 regulates macrophage infiltration and insulin sensitivity in mice fed a high-fat diet. J Endocrinol. 2015; 224:109–118.
Article13. Young Taek H. A concise synthesis of rhodanthpyrone A and B, natural 4-(hydroxyphenyl)-substituted a-pyrones. Nat Prod Commun. 2017; 12:95–98.14. Wang Z, Ka SO, Han YT, Bae EJ. Dihydropyranoaurone compound damaurone D inhibits LPS-induced inflammation and liver injury by inhibiting NF-κB and MAPK signaling independent of AMPK. Arch Pharm Res. 2018; 41:314–323.
Article15. Chawla A, Nguyen KD, Goh YP. Macrophage-mediated inflammation in metabolic disease. Nat Rev Immunol. 2011; 11:738–749.
Article16. Osborn O, Olefsky JM. The cellular and signaling networks linking the immune system and metabolism in disease. Nat Med. 2012; 18:363–374.
Article17. Russo L, Lumeng CN. Properties and functions of adipose tissue macrophages in obesity. Immunology. 2018; 155:407–417.
Article18. Olefsky JM, Glass CK. Macrophages, inflammation, and insulin resistance. Annu Rev Physiol. 2010; 72:219–246.
Article19. Chi H, Barry SP, Roth RJ, Wu JJ, Jones EA, Bennett AM, Flavell RA. Dynamic regulation of pro- and anti-inflammatory cytokines by MAPK phosphatase 1 (MKP-1) in innate immune responses. Proc Natl Acad Sci U S A. 2006; 103:2274–2279.
Article20. Kim HJ, Lee HS, Chong YH, Kang JL. p38 Mitogen-activated protein kinase up-regulates LPS-induced NF-kappaB activation in the development of lung injury and RAW 2647 macrophages. Toxicology. 2006; 225:36–47.21. Hayden MS, Ghosh S. Regulation of NF-κB by TNF family cytokines. Semin Immunol. 2014; 26:253–266.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Aloe-emodin inhibits Pam₃CSK₄-induced MAPK and NF-κB signaling through TLR2 in macrophages
- Beauvericin, a cyclic peptide, inhibits inflammatory responses in macrophages by inhibiting the NF-κB pathway
- Effect of Erythromycin on Pro-inflammatory Signalings by Particles
- Anti-Inflammatory Effect of Rosa rugosa Flower Extract in Lipopolysaccharide-Stimulated RAW264.7 Macrophages
- Anti-inflammatory effects of proanthocyanidin-rich red rice extract via suppression of MAPK, AP-1 and NF-κB pathways in Raw 264.7 macrophages
