Interaction of Wnt5a with Notch1 is Critical for the Pathogenesis of Psoriasis
- Affiliations
-
- 1Department of Dermatology, Asan Institute for Life Sciences, Asan Medical Center, University of Ulsan College of Medicine, Seoul, Korea. csesnumd@gmail.com
- 2Department of Dermatology, Hanyang University Hospital, Hanyang University College of Medicine, Seoul, Korea.
- 3Department of Dermatology, Chungnam National University College of Medicine, Daejeon, Korea.
- KMID: 2429481
- DOI: http://doi.org/10.5021/ad.2016.28.1.45
Abstract
- BACKGROUND
Psoriasis is characterized by uncontrolled hyperproliferation, aberrant differentiation, and dermal infiltration of immune cells. Recent studies have reported that Wnt5a and Notch1 signaling are altered in psoriatic skin lesions.
OBJECTIVE
We aimed to investigate the interaction of Wnt5a with Notch 1 with respect to inflammation-mediated epidermal hyperproliferation in psoriasis.
METHODS
Expression of Wnt5a and Notch1 signaling-related proteins were examined in psoriatic skin biopsies. Wnt5a was upregulated in human keratinocytes by treating the cells with its recombinant form (rWnt5a).
RESULTS
In psoriatic lesions, expression of Wnt5a increased while that of Notch1 decreased when compared to that in non-lesional and normal skin. Treatment with rWnt5a increased the proliferation of keratinocytes and increased their secretion of interleukin (IL)-23, IL-12, and tumor necrosis factor (TNF)-alpha. Further, exposure of keratinocytes to IL-1alpha, TNF-alpha, transforming growth factor-alpha, and interferon-gamma downregulated Notch1 as well as HES 1, which is downstream to Notch1, but increased the Wnt5a levels. The upregulated Wnt5a in keratinocytes downregulated both Notch1 and HES1.
CONCLUSION
Our data suggest that Wnt5a and Notch1 signaling exert counteracting influences on each other and are involved, in part, in the pathomechanism of psoriasis.
MeSH Terms
Figure
-
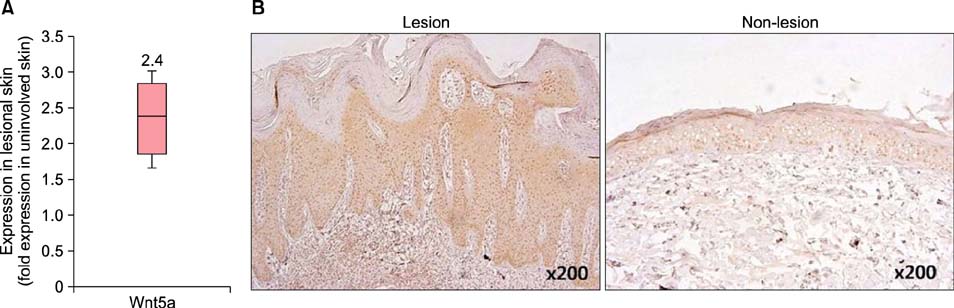
Fig. 1 Wnt5a expression in psoriasis. (A) Quantitative reverse transcription polymerase chain reaction results show that Wnt5a is upregulated approximately 2.4-fold in psoriatic lesional skin compared with that in non-lesional samples (n=10, paired samples). The box plot represents the log ratio of the expression levels in lesional and non-lesional psoriatic skin samples. Upper and lower borders of the boxes indicate 75th and 25th percentiles, respectively, whereas horizontal lines within boxes denote medians. (B) Immunohistochemical analyses show that Wnt5a is increased in all the layers of psoriatic lesions (left) compared with non-lesional skin (right). Samples were stained with DAB (3,3'-diaminobenzidine; brown color) and counterstained with hematoxylin (purple).

Fig. 2 Notch1 and hairy and enhancer of split (HES) 1 expression in psoriasis. (A) Quantitative reverse transcription polymerase chain reaction results did not yield any difference in the mRNA levels of NOTCH1 between psoriatic lesional and non-lesional skin samples, but revealed a decrease in HES1 mRNA expression in the former. (B) Immunohistochemical analyses (DAB [3,3'-diaminobenzidine] staining, ×400) showed that protein levels of Notch1 (upper panel) were decreased in all the layers of psoriatic lesions (left) compared with non-lesional skin samples (right). HES1 (lower panel) was also decreased at the protein level in psoriatic lesional samples.
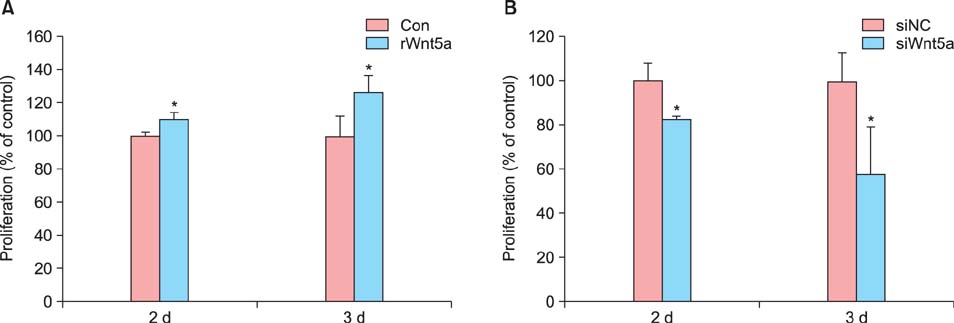
Fig. 3 The effects of Wnt5a on normal human keratinocyte (NHK) proliferation. NHKs were treated with (A) rWnt5a (100 ng/ml) and (B) siRNAs against Wnt5a for 2 and 3 d, and then examined by Cell Proliferation Bromodeoxyuridine (BrdU) enzyme-linked immunosorbent assay. The graph shows the percentage of BrdU-positive cells with respect to the control. Data represent the mean±standard deviation. Con: control, d: day. *p-value<0.05, determined by the Student's t-test.
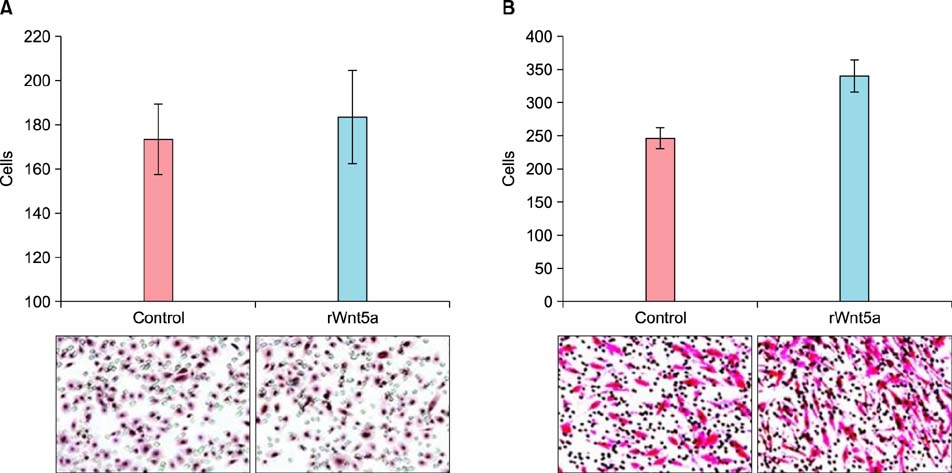
Fig. 4 Migration assays in neonatal normal human keratinocytes (NHKs) and human dermal fibroblasts (HDFs). NHKs (A) and HDFs (B) were cultured in either Wnt5a-conditioned or control media for 3 d, following which the cells (A: 1×106, B: 5×104) were incubated on transwell membranes (8-µm pore size) for 24 h. Migrated cells were stained with hematoxylin and eosin, and then enumerated under a microscope (×100).
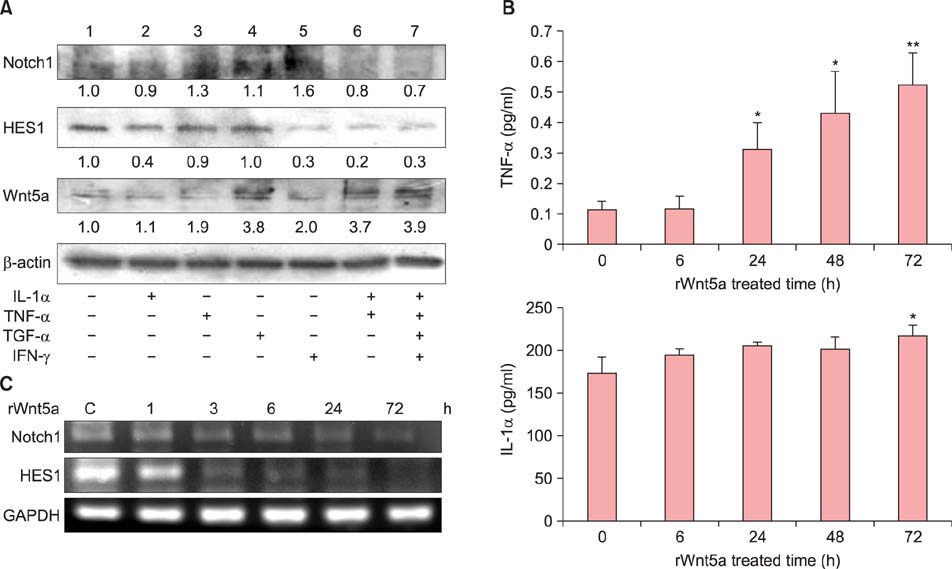
Fig. 5 Association of Wnt5a and Notch1 expression with the cytokine environment. (A) Neonatal normal human keratinocytes (NHKs) were stimulated with tumor necrosis factor (TNF)-α, interferon (IFN)-γ, transforming growth factor (TGF)-α, and interleukin (IL)-1α, either alone or with a cytokine mixture containing TNF-α, IFN-γ, TGF-α, and IL-1α for 24 h. Western blot analyses detected the expression of Notch1, hairy and enhancer of split (HES) 1, and Wnt5a. (1) Untreated NHKs (control), (2) IL-1α (10 ng/ml), (3) TNF-α (10 ng/ml), (4) TGF-α (24 ng/ml), (5) IFN-γ (20 ng/ml), (6) cytokine mixture of IL-1α (10 ng/ml), and TNF-α (10 ng/ml), (7) IL-1α (10 ng/ml), TNF-α (10 ng/ml), TGF-α (24 ng/ml), and IFN-γ (20 ng/ml)-treated NHKs. (B) Analysis of rWnt5a-treated NHK culture media by enzyme-linked immunosorbent assay (ELISA). Cells were treated with rWnt5a (100 ng/ml) for 6, 24, 48, and 72 h, following which the culture media were harvested. TNF-α and IL-1α production (pg/ml) were determined by ELISA. Data represent the mean±standard deviation. *p-value<0.05, **p-value<0.01; determined by the Student's t-test. (C) NHKs were treated with rWnt5a (100 ng/ml) for 1, 3, 6, 24, and 72 h, following which the expression levels of NOTCH1 and HES1 were determined by reverse transcription polymerase chain reaction.
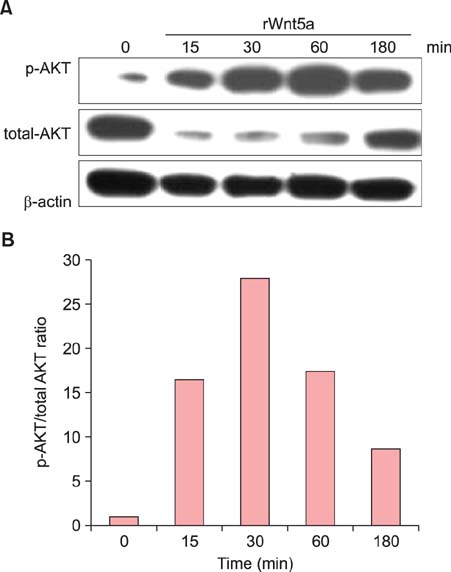
Fig. 6 Phospho (p)-AKT signaling in rWnt5a-treated neonatal normal human keratinocytes (NHKs). NHKs, serum-deprived for 24 h, were either treated with 100 ng/ml of rWnt5a or left untreated, and harvested after 15 min, 30 min, 1 h, and 3 h. Proteins in the cell lysates were resolved by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), and analyzed by western blotting using antibodies against p-AKT and total AKT. (A) Representative western blot for total AKT and p-AKT, showing an increase in the phosphorylated (activated) form of AKT in rWnt5a-treated NHKs. (B) Intensities of p-AKT and total AKT signals were determined using the BioRad VersaDoc Imaging System. The p-AKT/total AKT ratio is indicated for each time point.
Reference
-
1. Nestle FO, Kaplan DH, Barker J. Psoriasis. N Engl J Med. 2009; 361:496–509.
Article2. Reischl J, Schwenke S, Beekman JM, Mrowietz U, Stürzebecher S, Heubach JF. Increased expression of Wnt5a in psoriatic plaques. J Invest Dermatol. 2007; 127:163–169.
Article3. Romanowska M, Evans A, Kellock D, Bray SE, McLean K, Donandt S, et al. Wnt5a exhibits layer-specific expression in adult skin, is upregulated in psoriasis, and synergizes with type 1 interferon. PLoS One. 2009; 4:e5354.
Article4. Gudjonsson JE, Johnston A, Stoll SW, Riblett MB, Xing X, Kochkodan JJ, et al. Evidence for altered Wnt signaling in psoriatic skin. J Invest Dermatol. 2010; 130:1849–1859.
Article5. Nicolas M, Wolfer A, Raj K, Kummer JA, Mill P, van Noort M, et al. Notch1 functions as a tumor suppressor in mouse skin. Nat Genet. 2003; 33:416–421.
Article6. Devgan V, Mammucari C, Millar SE, Brisken C, Dotto GP. p21WAF1/Cip1 is a negative transcriptional regulator of Wnt4 expression downstream of Notch1 activation. Genes Dev. 2005; 19:1485–1495.
Article7. Cheng CW, Yeh JC, Fan TP, Smith SK, Charnock-Jones DS. Wnt5a-mediated non-canonical Wnt signalling regulates human endothelial cell proliferation and migration. Biochem Biophys Res Commun. 2008; 365:285–290.
Article8. Masckauchán TN, Agalliu D, Vorontchikhina M, Ahn A, Parmalee NL, Li CM, et al. Wnt5a signaling induces proliferation and survival of endothelial cells in vitro and expression of MMP-1 and Tie-2. Mol Biol Cell. 2006; 17:5163–5172.
Article9. Yu JM, Jun ES, Jung JS, Suh SY, Han JY, Kim JY, et al. Role of Wnt5a in the proliferation of human glioblastoma cells. Cancer Lett. 2007; 257:172–181.
Article10. Kawasaki A, Torii K, Yamashita Y, Nishizawa K, Kanekura K, Katada M, et al. Wnt5a promotes adhesion of human dermal fibroblasts by triggering a phosphatidylinositol-3 kinase/Akt signal. Cell Signal. 2007; 19:2498–2506.
Article11. Säfholm A, Leandersson K, Dejmek J, Nielsen CK, Villoutreix BO, Andersson T. A formylated hexapeptide ligand mimics the ability of Wnt-5a to impair migration of human breast epithelial cells. J Biol Chem. 2006; 281:2740–2749.
Article12. Blumenthal A, Ehlers S, Lauber J, Buer J, Lange C, Goldmann T, et al. The Wingless homolog WNT5A and its receptor Frizzled-5 regulate inflammatory responses of human mononuclear cells induced by microbial stimulation. Blood. 2006; 108:965–973.
Article13. Pereira C, Schaer DJ, Bachli EB, Kurrer MO, Schoedon G. Wnt5A/CaMKII signaling contributes to the inflammatory response of macrophages and is a target for the antiinflammatory action of activated protein C and interleukin-10. Arterioscler Thromb Vasc Biol. 2008; 28:504–510.
Article14. Mammucari C, Tommasi di Vignano A, Sharov AA, Neilson J, Havrda MC, Roop DR, et al. Integration of Notch 1 and calcineurin/NFAT signaling pathways in keratinocyte growth and differentiation control. Dev Cell. 2005; 8:665–676.
Article15. Nguyen BC, Lefort K, Mandinova A, Antonini D, Devgan V, Della Gatta G, et al. Cross-regulation between Notch and p63 in keratinocyte commitment to differentiation. Genes Dev. 2006; 20:1028–1042.
Article16. Lai EC. Notch signaling: control of cell communication and cell fate. Development. 2004; 131:965–973.
Article17. Okuyama R, Tagami H, Aiba S. Notch signaling: its role in epidermal homeostasis and in the pathogenesis of skin diseases. J Dermatol Sci. 2008; 49:187–194.
Article18. Thélu J, Rossio P, Favier B. Notch signalling is linked to epidermal cell differentiation level in basal cell carcinoma, psoriasis and wound healing. BMC Dermatol. 2002; 2:7.
Article19. Iizuka H, Takahashi H, Honma M, Ishida-Yamamoto A. Unique keratinization process in psoriasis: late differentiation markers are abolished because of the premature cell death. J Dermatol. 2004; 31:271–276.
Article20. Okuyama R, Ogawa E, Nagoshi H, Yabuki M, Kurihara A, Terui T, et al. p53 homologue, p51/p63, maintains the immaturity of keratinocyte stem cells by inhibiting Notch1 activity. Oncogene. 2007; 26:4478–4488.
Article21. Rangarajan A, Talora C, Okuyama R, Nicolas M, Mammucari C, Oh H, et al. Notch signaling is a direct determinant of keratinocyte growth arrest and entry into differentiation. EMBO J. 2001; 20:3427–3436.
Article22. Blaumueller CM, Qi H, Zagouras P, Artavanis-Tsakonas S. Intracellular cleavage of Notch leads to a heterodimeric receptor on the plasma membrane. Cell. 1997; 90:281–291.
Article23. Kim JE, Lee JH, Jeong KH, Kim GM, Kang H. Notch intracellular domain expression in various skin fibroproliferative diseases. Ann Dermatol. 2014; 26:332–337.
Article24. Collins BJ, Kleeberger W, Ball DW. Notch in lung development and lung cancer. Semin Cancer Biol. 2004; 14:357–364.
Article25. Okuyama R, Nguyen BC, Talora C, Ogawa E, Tommasi di Vignano A, Lioumi M, et al. High commitment of embryonic keratinocytes to terminal differentiation through a Notch1-caspase 3 regulatory mechanism. Dev Cell. 2004; 6:551–562.
Article26. Lefort K, Mandinova A, Ostano P, Kolev V, Calpini V, Kolfschoten I, et al. Notch1 is a p53 target gene involved in human keratinocyte tumor suppression through negative regulation of ROCK1/2 and MRCKalpha kinases. Genes Dev. 2007; 21:562–577.
Article27. Nickoloff BJ, Qin JZ, Chaturvedi V, Denning MF, Bonish B, Miele L. Jagged-1 mediated activation of notch signaling induces complete maturation of human keratinocytes through NF-kappaB and PPARgamma. Cell Death Differ. 2002; 9:842–855.
Article28. Ota T, Takekoshi S, Takagi T, Kitatani K, Toriumi K, Kojima T, et al. Notch signaling may be involved in the abnormal differentiation of epidermal keratinocytes in psoriasis. Acta Histochem Cytochem. 2014; 47:175–183.
Article29. Rooney P, Connolly M, Gao W, McCormick J, Biniecka M, Sullivan O, et al. Notch-1 mediates endothelial cell activation and invasion in psoriasis. Exp Dermatol. 2014; 23:113–118.
Article30. Zhou X, Krueger JG, Kao MC, Lee E, Du F, Menter A, et al. Novel mechanisms of T-cell and dendritic cell activation revealed by profiling of psoriasis on the 63,100-element oligonucleotide array. Physiol Genomics. 2003; 13:69–78.
Article31. van Amerongen R, Mikels A, Nusse R. Alternative wnt signaling is initiated by distinct receptors. Sci Signal. 2008; 1:re9.
Article32. Couso JP, Martinez Arias A. Notch is required for wingless signaling in the epidermis of Drosophila. Cell. 1994; 79:259–272.
Article33. Ayyanan A, Civenni G, Ciarloni L, Morel C, Mueller N, Lefort K, et al. Increased Wnt signaling triggers oncogenic conversion of human breast epithelial cells by a Notch-dependent mechanism. Proc Natl Acad Sci U S A. 2006; 103:3799–3804.
Article34. Ann EJ, Kim HY, Seo MS, Mo JS, Kim MY, Yoon JH, et al. Wnt5a controls Notch1 signaling through CaMKII-mediated degradation of the SMRT corepressor protein. J Biol Chem. 2012; 287:36814–36829.
Article35. Katoh M, Katoh M. Transcriptional mechanisms of WNT5A based on NF-kappaB, Hedgehog, TGFbeta, and Notch signaling cascades. Int J Mol Med. 2009; 23:763–769.36. Koyanagi M, Bushoven P, Iwasaki M, Urbich C, Zeiher AM, Dimmeler S. Notch signaling contributes to the expression of cardiac markers in human circulating progenitor cells. Circ Res. 2007; 101:1139–1145.
Article37. Zhu X, Wu Y, Huang S, Chen Y, Tao Y, Wang Y, et al. Overexpression of Wnt5a in mouse epidermis causes no psoriasis phenotype but an impairment of hair follicle anagen development. Exp Dermatol. 2014; 23:926–928.
Article38. Kim JE, Won CH, Bak H, Kositratna G, Manstein D, Dotto GP, et al. Gene profiling analysis of the early effects of ablative fractional carbon dioxide laser treatment on human skin. Dermatol Surg. 2013; 39:1033–1043.
Article39. Suárez-Fariñas M, Fuentes-Duculan J, Lowes MA, Krueger JG. Resolved psoriasis lesions retain expression of a subset of disease-related genes. J Invest Dermatol. 2011; 131:391–400.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- A Case showing Improved Psoriasis along the Hemiparetic area after Cerebrovascular Accident
- Potential biomarkers and signaling pathways associated with the pathogenesis of primary salivary gland carcinoma: a bioinformatics study
- Coexistence of Bullous Pemphigoid and Psoriasis: A Case Report and Review of the Literature
- Significance of Notch Expression in Acute Myeloid Leukemia
- Immunohistochemical studies of Interleukin-8 and Tumor necrosis factor-alpha on psoriatic lesions
