Chicken RNA-binding protein T-cell internal antigen-1 contributes to stress granule formation in chicken cells and tissues
- Affiliations
-
- 1Department of Avian Infectious Diseases, Shanghai Veterinary Research Institute, Chinese Academy of Agricultural Science, Shanghai 200241, China. shoveldeen@shvri.ac.cn
- 2College of Animal Science and Technology, Shandong Agricultural University, Taian 271018, China.
- 3College of Animal Science and Technology, Jilin Agricultural University, Changchun 130118, China.
- 4Jiangsu Co-innovation Center for Prevention and Control of Important Animal Infectious Diseases and Zoonoses, Yangzhou 225009, China.
- KMID: 2402689
- DOI: http://doi.org/10.4142/jvs.2018.19.1.3
Abstract
- T-cell internal antigen-1 (TIA-1) has roles in regulating alternative pre-mRNA splicing, mRNA translation, and stress granule (SG) formation in human cells. As an evolutionarily conserved response to environmental stress, SGs have been reported in various species. However, SG formation in chicken cells and the role of chicken TIA-1 (cTIA-1) in SG assembly has not been elucidated. In the present study, we cloned cTIA-1 and showed that it facilitates the assembly of canonical SGs in both human and chicken cells. Overexpression of the chicken prion-related domain (cPRD) of cTIA-1 that bore an N-terminal green fluorescent protein (GFP) tag (pntGFP-cPRD) or Flag tag (pFlag-cPRD) induced the production of typical SGs. However, C-terminal GFP-tagged cPRD induced notably large cytoplasmic granules that were devoid of endogenous G3BP1 and remained stable when exposed to cycloheximide, indicating that these were not typical SGs, and that the pntGFP tag influences cPRD localization. Finally, endogenous cTIA-1 was recruited to SGs in chicken cells and tissues under environmental stress. Taken together, our study provide evidence that cTIA-1 has a role in canonical SG formation in chicken cells and tissues. Our results also indicate that cPRD is necessary for SG aggregation.
MeSH Terms
Figure
-
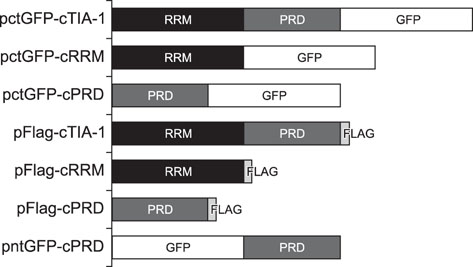
Fig. 1 Schematic representation of 7 expression plasmids, pctGFP-cTIA-1, pctGFP-cRRM, pctGFP-cPRD, pFlag-cTIA-1, pFlag-cRRM, pFlag-cPRD, and pntGFP-cPRD. GFP, green fluorescent protein; cTIA-1, chicken T-cell internal antigen-1; RRM, RNA recognition motif; PRD, prion-related domain; cRRM, chicken RRM; cPRD, chicken PRD.
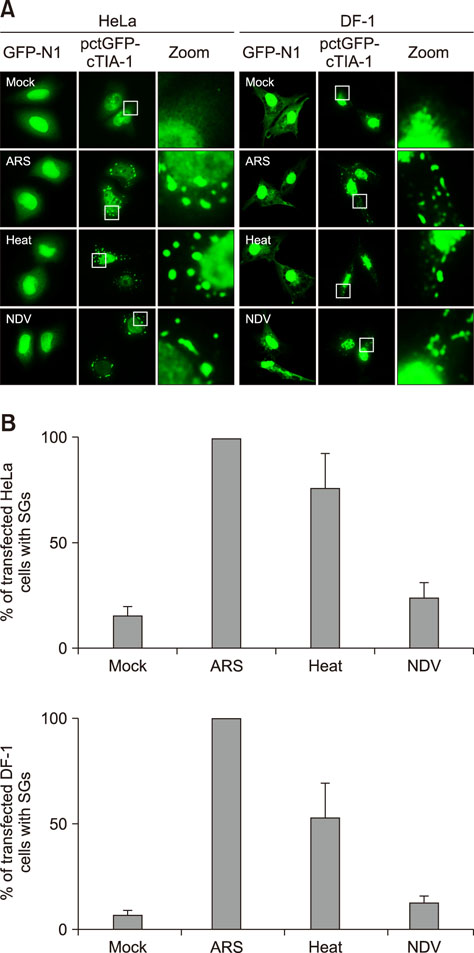
Fig. 2 Cytoplasmic fluorescent dots were detected in pctGFP-cTIA-1-transfected HeLa and DF-1 cells that were exposed to environmental stressors. (A) HeLa and DF-1 cells were transfected with pEGFP-N1 and pctGFP-cTIA-1, respectively, followed by treatment at 24 h post-transfection with mock treatment, treatment with arsenite (ARS), heat shock, or infection by Newcastle disease virus (NDV) at an multiplicity of infection of 1 for 18 h. The cells were collected, fixed, and visualized by confocal microscopy. (B) The percentages of cells containing stress granule (SGs) per high-powered fields (HPF) were quantified visually by manual counting. A total of 10 pictures in 20 HPF were randomly captured in different fields. The data are expressed as the mean of three independent experiments. GFP, green fluorescent protein; cTIA-1, chicken T-cell internal antigen-1.
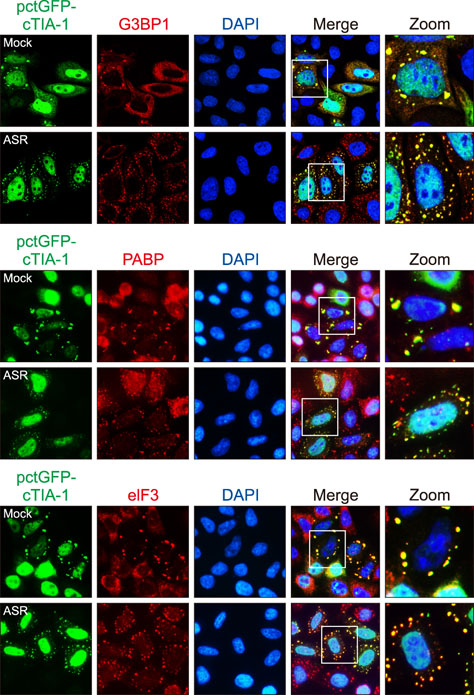
Fig. 3 Exogenously expressed pctGFP-cTIA-1 localized with endogenous stress granule markers in HeLa cells. HeLa cells transfected with pctGFP-cTIA-1 were mock treated or treated with arsenite (ARS) for 30 min. The cells were fixed and stained with an anti-G3BP1, PABP, or eIF3 antibody, respectively, and then visualized by confocal microscopy. GFP, green fluorescent protein; cTIA-1, chicken T-cell internal antigen-1; G3BP1, GTPase-activating protein-binding protein 1; DAPI, 4′, 6-diamidino-2-phenylindole; PABP, poly(A)-binding protein; eIF3, eukaryotic initiation factor 3.

Fig. 4 pctGFP-cPRD induced the formation of atypical large granules that were stable under cycloheximide (CHX) treatment. (A) HeLa cells transfected with pctGFP-cTIA-1, pctGFP-cPRD, or pctGFP-cRRM were mock treated or treated with arsenite (ARS) for 30 min. The cells were fixed and stained with an anti-G3BP1 antibody and then visualized by confocal microscopy. (B) HeLa cells transfected with pctGFP-cTIA-1, pctGFP-cPRD, or pctGFP-cRRM were treated with CHX. After 2 h, the cells were mock treated or treated with ARS for 30 min. The cells were fixed and stained with anti-G3BP1 antibody and then visualized by confocal microscopy. G3BP1, GTPase-activating protein-binding protein 1; DAPI, 4′, 6-diamidino-2-phenylindole; GFP, green fluorescent protein; cTIA-1, chicken T-cell internal antigen-1; cPRD, chicken prion-related domain; cRRM, chicken RNA recognition motif.
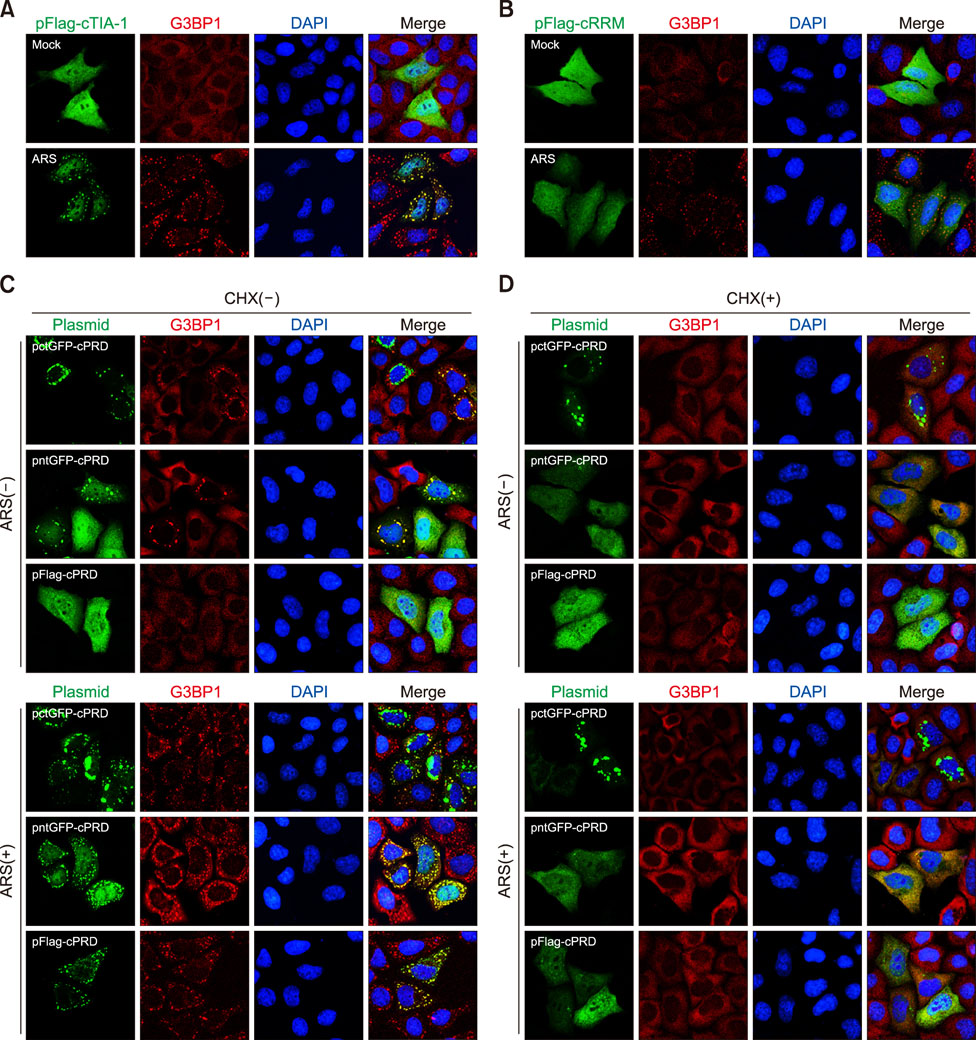
Fig. 5 The effect of the GFP tag on cPRD localization. HeLa cells transfected with pFlag-cTIA-1 (A) or pFlag-cRRM (B) were mock treated or treated with arsenite (ARS) for 30 min. Cells were fixed and stained with anti-Flag (only for pFlag-cTIA-1 transfection group) and anti-G3BP1 antibody and then visualized by confocal microscopy. (C) HeLa cells transfected with pctGFP-cPRD, pntGFP-cPRD, or pFlag-cPRD were mock treated or treated with ARS for 30 min. Cells were fixed and stained with anti-Flag (only for pFlag-cTIA-1 transfection group) and anti-G3BP1 antibody and then visualized by confocal microscopy. (D) HeLa cells transfected with pctGFP-cPRD, pntGFP-cPRD, or pFlag-cPRD were treated with cycloheximide (CHX). After 2 h, the cells were mock treated or treated with ARS for 30 min. The cells were fixed and stained with an anti-G3BP1 antibody and then visualized by confocal microscopy. cTIA-1, chicken T-cell internal antigen-1; G3BP1, GTPase-activating protein-binding protein 1; DAPI, 4′, 6-diamidino-2-phenylindole; cRRM, chicken RNA recognition motif; GFP, green fluorescent protein; cPRD, chicken prion-related domain.
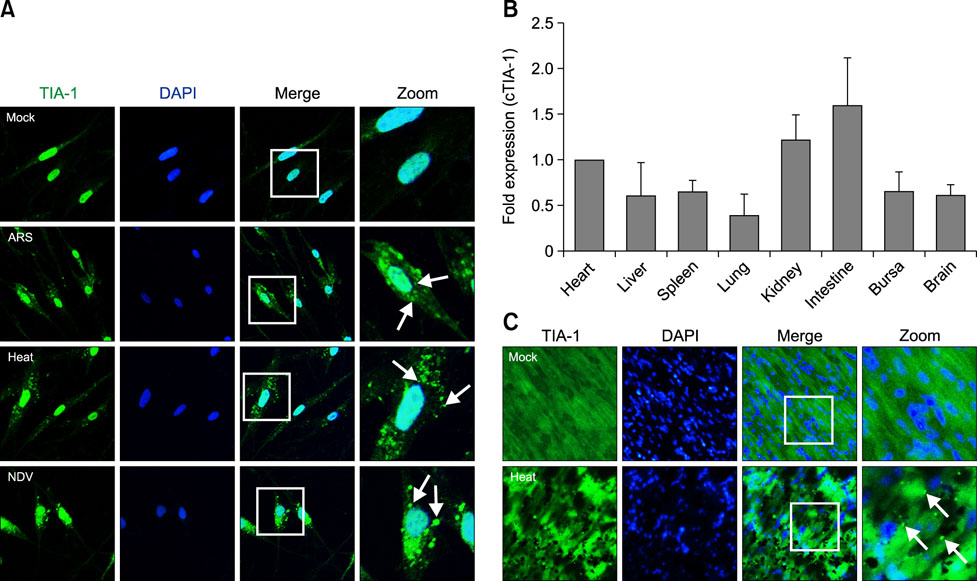
Fig. 6 (A) HeLa cells were mock treated or treated with arsenite (ARS), heat shock, or infected with a virus. The cells were fixed and stained with an anti-TIA-1 antibody and then visualized by confocal microscopy. Arrows, stress granule (SG) dots. (B) The transcription levels of cTIA-1 were detected by performing real-time polymerase chain reaction. Fold-expressions were calculated based on the level of the cTIA-1 gene in the heart. (C) Two groups of three 7-day-old specific-pathogen-free chickens were mock treated or treated with heat shock. At the indicated time, the mock- and heat-shock-treated chicken hearts were dissected, sectioned, and stained with anti-TIA-1 antibody and then visualized by confocal microscopy. Arrows, SG dots. TIA-1, T-cell internal antigen-1; DAPI, 4′, 6-diamidino-2-phenylindole; NDV, Newcastle disease virus; cTIA-1, chicken TIA-1.
Reference
-
1. Anderson P, Kedersha N. RNA granules. J Cell Biol. 2006; 172:803–808.
Article2. Buchan JR, Muhlrad D, Parker R. P bodies promote stress granule assembly in Saccharomyces cerevisiae. J Cell Biol. 2008; 183:441–455.3. Collier NC, Schlesinger MJ. The dynamic state of heat shock proteins in chicken embryo fibroblasts. J Cell Biol. 1986; 103:1495–1507.
Article4. Cubitt AB, Heim R, Adams SR, Boyd AE, Gross LA, Tsien RY. Understanding, improving and using green fluorescent proteins. Trends Biochem Sci. 1995; 20:448–455.
Article5. De Maio A. Heat shock proteins: facts, thoughts, and dreams. Shock. 1999; 11:1–12.
Article6. Efthymiou CA, Mocanu MM, de Belleroche J, Wells DJ, Latchmann DS, Yellon DM. Heat shock protein 27 protects the heart against myocardial infarction. Basic Res Cardiol. 2004; 99:392–394.
Article7. Förch P, Puig O, Kedersha N, Martínez C, Granneman S, Séraphin B, Anderson P, Valcárcel J. The apoptosis-promoting factor TIA-1 is a regulator of alternative pre-mRNA splicing. Mol Cell. 2000; 6:1089–1098.
Article8. Gallouzi IE, Brennan CM, Stenberg MG, Swanson MS, Eversole A, Maizels N, Steitz JA. HuR binding to cytoplasmic mRNA is perturbed by heat shock. Proc Natl Acad Sci U S A. 2000; 97:3073–3078.
Article9. Gerdes HH, Kaether C. Green fluorescent protein: applications in cell biology. FEBS Lett. 1996; 389:44–47.
Article10. Gilks N, Kedersha N, Ayodele M, Shen L, Stoecklin G, Dember LM, Anderson P. Stress granule assembly is mediated by prion-like aggregation of TIA-1. Mol Biol Cell. 2004; 15:5383–5398.
Article11. Kandasamy K, Joseph K, Subramaniam K, Raymond JR, Tholanikunnel BG. Translational control of beta2-adrenergic receptor mRNA by T-cell-restricted intracellular antigen-related protein. J Biol Chem. 2005; 280:1931–1943.
Article12. Kedersha N, Anderson P. Stress granules: sites of mRNA triage that regulate mRNA stability and translatability. Biochem Soc Trans. 2002; 30:963–969.
Article13. Kedersha N, Chen S, Gilks N, Li W, Miller IJ, Stahl J, Anderson P. Evidence that ternary complex (eIF2-GTP-tRNAiMet)-deficient preinitiation complexes are core constituents of mammalian stress granules. Mol Biol Cell. 2002; 13:195–210.
Article14. Kedersha N, Cho MR, Li W, Yacono PW, Chen S, Gilks N, Golan DE, Anderson P. Dynamic shuttling of TIA-1 accompanies the recruitment of mRNA to mammalian stress granules. J Cell Biol. 2000; 151:1257–1268.
Article15. Kedersha NL, Gupta M, Li W, Miller I, Anderson P. RNA-binding proteins TIA-1 and TIAR link the phosphorylation of eIF-2 alpha to the assembly of mammalian stress granules. J Cell Biol. 1999; 147:1431–1442.
Article16. Le Guiner C, Gesnel MC, Breathnach R. TIA-1 or TIAR is required for DT40 cell viability. J Biol Chem. 2003; 278:10465–10476.
Article17. Mangiardi DA, McLaughlin-Williamson K, May KE, Messana EP, Mountain DC, Cotanche DA. Progression of hair cell ejection and molecular markers of apoptosis in the avian cochlea following gentamicin treatment. J Comp Neurol. 2004; 475:1–18.
Article18. Nover L, Scharf KD, Neumann D. Formation of cytoplasmic heat shock granules in tomato cell cultures and leaves. Mol Cell Biol. 1983; 3:1648–1655.
Article19. Ooie T, Takahashi N, Saikawa T, Nawata T, Arikawa M, Yamanaka K, Hara M, Shimada T, Sakata T. Single oral dose of geranylgeranylacetone induces heat-shock protein 72 and renders protection against ischemia/reperfusion injury in rat heart. Circulation. 2001; 104:1837–1843.
Article20. Piecyk M, Wax S, Beck AR, Kedersha N, Gupta M, Maritim B, Chen S, Gueydan C, Kruys V, Streuli M, Anderson P. TIA-1 is a translational silencer that selectively regulates the expression of TNF-alpha. EMBO J. 2000; 19:4154–4163.
Article21. Piotrowska J, Hansen SJ, Park N, Jamka K, Sarnow P, Gustin KE. Stable formation of compositionally unique stress granules in virus-infected cells. J Virol. 2010; 84:3654–3665.
Article22. Serio TR, Lindquist SL. [PSI+], SUP35, and chaperones. Adv Protein Chem. 2001; 57:335–366.
Article23. Sun Y, Ding N, Ding SS, Yu S, Meng C, Chen H, Qiu X, Zhang S, Yu Y, Zhan Y, Ding C. Goose RIG-I functions in innate immunity against Newcastle disease virus infections. Mol Immunol. 2013; 53:321–327.
Article24. Sun Y, Yu S, Ding N, Meng C, Meng S, Zhang S, Zhan Y, Qiu X, Tan L, Chen H, Song C, Ding C. Autophagy benefits the replication of Newcastle disease virus in chicken cells and tissues. J Virol. 2014; 88:525–537.
Article25. Tourrière H, Chebli K, Zekri L, Courselaud B, Blanchard JM, Bertrand E, Tazi J. The RasGAP-associated endoribonuclease G3BP assembles stress granules. J Cell Biol. 2003; 160:823–831.
Article26. Vaheri A, Ruoslahti E, Sarvas M, Nurminen M. Mitogenic effect by lipopolysaccharide and pokeweed lectin on density-inhibited chick embryo fibroblasts. J Exp Med. 1973; 138:1356–1364.
Article27. Zhang T, Delestienne N, Huez G, Kruys V, Gueydan C. Identification of the sequence determinants mediating the nucleo-cytoplasmic shuttling of TIAR and TIA-1 RNA-binding proteins. J Cell Sci. 2005; 118:5453–5463.
Article28. Zhu H, Hasman RA, Young KM, Kedersha NL, Lou H. U1 snRNP-dependent function of TIAR in the regulation of alternative RNA processing of the human calcitonin/CGRP pre-mRNA. Mol Cell Biol. 2003; 23:5959–5971.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Neuronal RNA granule contains ApCPEB1, a novel cytoplasmic polyadenylation element binding protein, in Aplysia sensory neuron
- Downregulation of cellular prion protein inhibited the proliferation and invasion and induced apoptosis of Marek's disease virus-transformed avian T cells
- Isolation and Characterization of Chicken NPAS3
- (3H)Ryanodine binding sites of SR vesicles of the chicken pectoral muscle
- Isolation and identification of Cryptosporidium from various animals in Korea. III. Identification of Cryptosporidium baileyi from Korean chicken
