Serosal Cavities Contain Two Populations of Innate-like integrin α4highCD4+ T Cells, Integrin α4β1+α6β1+α4β7− and α4β1+α6β1−α4β7+ Cells
- Affiliations
-
- 1Division of Immunobiology, Department of Molecular Cell Biology, Sungkyunkwan University School of Medicine, Suwon 16419, Korea. tjkim@skku.edu
- 2Department of Surgery, Seoul National University College of Medicine, Seoul 03080, Korea.
- 3Department of Health Sciences and Technology, Samsung Advanced institute of health Sciences and Technology (SAIHST) and Samsung Medical Center, Sungkyunkwan University, Seoul 06351, Korea.
- KMID: 2400625
- DOI: http://doi.org/10.4110/in.2017.17.6.392
Abstract
- We previously reported peritoneal innate-like integrin α4 (CD49d)highCD4+ T cells that provided help for B-1a cells. Here we analyzed the expression of various integrin chains on the peritoneal and pleural integrin α4highCD4+ T cells and investigated the functional heterogeneity of the subpopulations based on the integrin expression. Pleural cavity contained a lower ratio of integrin α4highCD4+ T cells to integrin α4lowCD4+ T cells than peritoneal cavity, but the pleural integrin α4highCD4+ T cells have the same characteristics of the peritoneal integrin α4highCD4+ T cells. Most of integrin α4highCD4+ T cells were integrin β1highβ7−, but a minor population of integrin α4highCD4+ T cells was integrin β1+β7+. Interestingly, the integrin α4highβ1highβ7− CD4+ T cells expressed high levels of integrin α4β1 and α6β1, whereas integrin α4highβ1+β7+ CD4+ T cells expressed high levels of integrin α4β1 and α4β7, suggesting an alternative expression of integrin α6β1 or α4β7 in combination with α4β1 in respective major and minor populations of integrin α4highCD4+ T cells. The minor population, integrin α4highβ1+β7+ CD4+ T cells, were different from the integrin α4highβ1highβ7− CD4+ T cells in that they secreted a smaller amount of Th1 cytokines upon stimulation and expressed lower levels of Th1-related chemokine receptors CCR5 and CXCR3 than the integrin α4highβ1 highβ7− CD4+ T cells. In summary, the innate-like integrin α4highCD4+ T cells could be divided into 2 populations, integrin α4β1+α6β1+α4β7− and α4β1+α6β1−α4β7+ cells. The functional significance of serosal integrin α4β7+ CD4+ T cells needed to be investigated especially in view of mucosal immunity.
Keyword
MeSH Terms
Figure
-
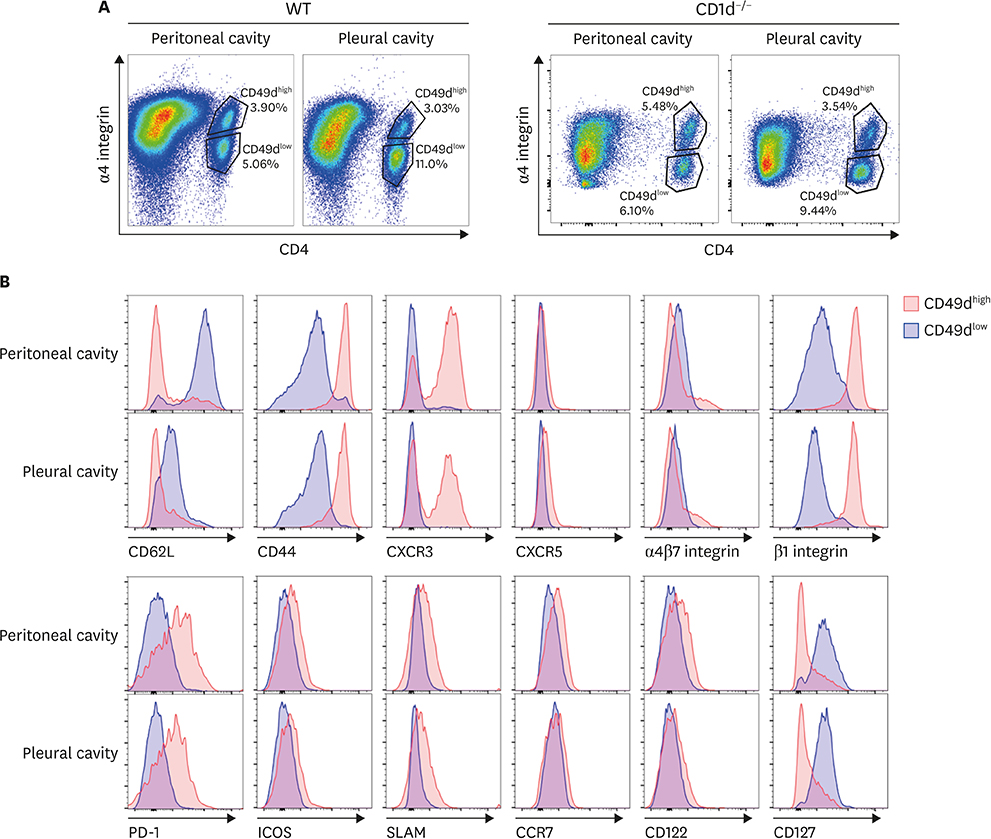
Figure 1 Expression of activation-related cell surface molecules in peritoneal and pleural integrin α4highCD4+ T cells. (A) PEC and PLC cells obtained from 8-week-old C57BL/6 WT or Cd1d −/− mice were examined for the expression of CD4 and integrin α4 (CD49d). (B) Expression of given cell surface proteins in gated peritoneal and pleural integrin α4highCD4+ T cells (red line, CD49dhigh) or CD49dlow T cells (blue line, CD49dlow). Data are representative of 12 separate experiments. WT, wild type.

Figure 2 Serosal integrin α4highCD4+ T cells are divided into integrin α4highβ1highCD4+ T and α4highβ1+β7+CD4+ T cells. (A) PEC and PLC CD4+ T cells or gated integrin α4highCD4+ T cells obtained from 8-week-old C57BL/6 wild type mice were shown for the expression of integrin β1 and β7. (B) Illustration for integrin combinations of integrin α4, β1, and β7 chains. (C) Cell surface expression of various integrin α chains in integrin α4highβ1highCD4+ T cells (red line, α4highβ1high), integrin α4highβ1+β7+CD4+ T cells (blue line, α4highβ1+β7+), and integrin α4lowCD4+ T cells (green line, α4low). Data are representative of at least 3 separate experiments.
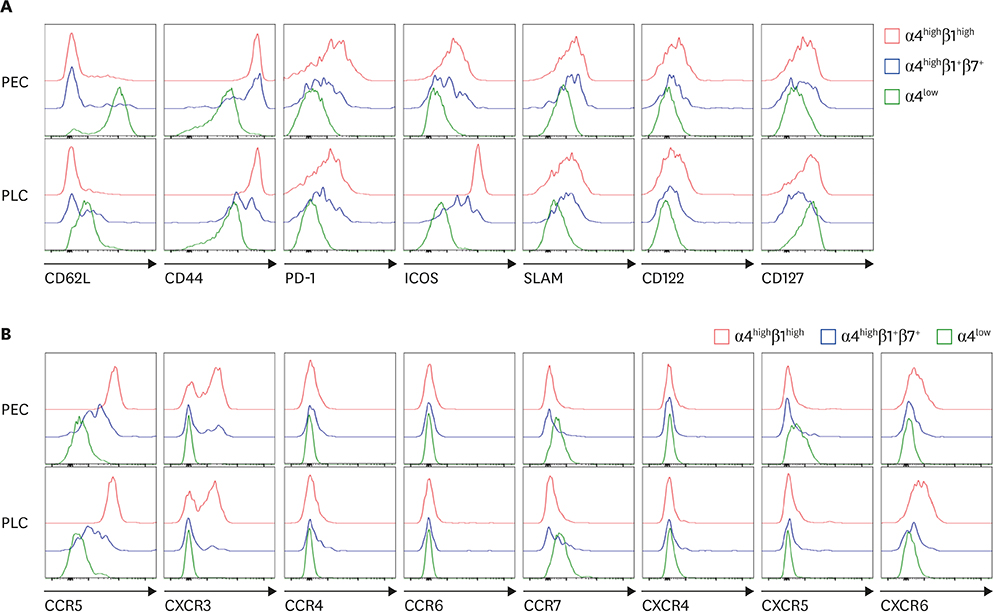
Figure 3 Comparison of various activation-related molecules (A) and chemokine receptors (B) among integrin α4highβ1highCD4+ T cells (red line), integrin α4highβ1+β7+CD4+ T cells (blue line), and integrin α4lowCD4+ T cells (green line). PEC and PLC cells obtained from 8-week-old C57BL/6 mice were examined for the expression of indicated cell surface proteins. Data are representative of 12 separate experiments.
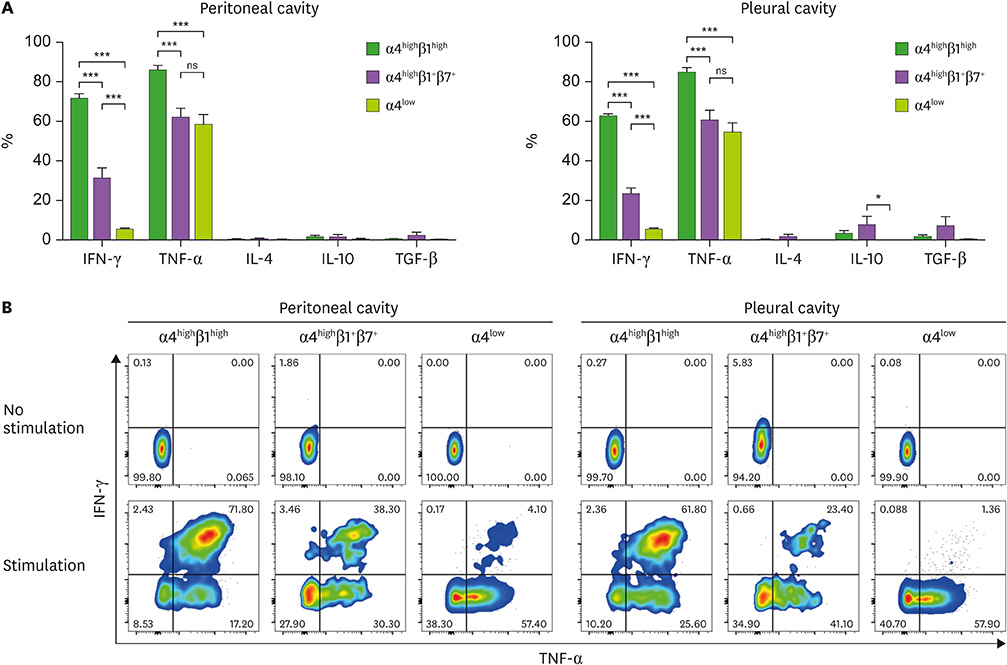
Figure 4 Cytokine production by integrin α4highβ1highCD4+ T cells, integrin α4highβ1+β7+CD4+ T cells, and integrin α4lowCD4+ T cells. (A, B) Peritoneal and pleural cavity cells harvested from 8-week-old C57BL/6 mice were stimulated in vitro with 50 ng/ml PMA and 1.5 mM ionomycin for 4 h. (A) Individual bars represent the percentages of given cytokine-producing cells among α4highβ1highCD4+ T cells (green, α4highβ1high), α4highβ1+β7+CD4+ T cells (purple, α4highβ1+β7+), and α4lowCD4+ T cells (yellowish green, α4low), detected by intracytoplasmic staining of given cytokines. (B) Representative flow cytometric data for given cytokines are shown with or without stimulation. Data are representative of 12 separate experiments. ns, not significant. *P<0.05; ***P<0.001.
Cited by 1 articles
-
Expansion and Sub-Classification of T Cell-Dependent Antibody Responses to Encompass the Role of Innate-Like T Cells in Antibody Responses
Chanho Park, Tae Jin Kim
Immune Netw. 2018;18(5):. doi: 10.4110/in.2018.18.e34.
Reference
-
1. Haas KM. B-1 lymphocytes in mice and nonhuman primates. Ann N Y Acad Sci. 2015; 1362:98–109.
Article2. Ganshina IV. Serous cavities of coelomic origin as possible organs of the immune system. Part 1. Biol Bull Rev. 2016; 6:497–504.
Article3. Moon H, Park C, Lee JG, Shin SH, Lee JH, Kho I, Kang K, Cha HS, Kim TJ. Early development in the peritoneal cavity of CD49dhigh Th1 memory phenotype CD4+ T cells with enhanced B cell helper activity. J Immunol. 2015; 195:564–575.
Article4. Weber GF, Chousterman BG, Hilgendorf I, Robbins CS, Theurl I, Gerhardt LM, Iwamoto Y, Quach TD, Ali M, Chen JW, et al. Pleural innate response activator B cells protect against pneumonia via a GM-CSF-IgM axis. J Exp Med. 2014; 211:1243–1256.
Article5. Rose DM, Han J, Ginsberg MH. Alpha4 integrins and the immune response. Immunol Rev. 2002; 186:118–124.6. Woodland DL, Kohlmeier JE. Migration, maintenance and recall of memory T cells in peripheral tissues. Nat Rev Immunol. 2009; 9:153–161.
Article7. Kadioglu A, De Filippo K, Bangert M, Fernandes VE, Richards L, Jones K, Andrew PW, Hogg N. The integrins Mac-1 and alpha4beta1 perform crucial roles in neutrophil and T cell recruitment to lungs during Streptococcus pneumoniae infection. J Immunol. 2011; 186:5907–5915.
Article8. Rott LS, Briskin MJ, Andrew DP, Berg EL, Butcher EC. A fundamental subdivision of circulating lymphocytes defined by adhesion to mucosal addressin cell adhesion molecule-1. Comparison with vascular cell adhesion molecule-1 and correlation with beta 7 integrins and memory differentiation. J Immunol. 1996; 156:3727–3736.9. Kurmaeva E, Lord JD, Zhang S, Bao JR, Kevil CG, Grisham MB, Ostanin DV. T cell-associated α4β7 but not α4β1 integrin is required for the induction and perpetuation of chronic colitis. Mucosal Immunol. 2014; 7:1354–1365.
Article10. Millward JM, Løbner M, Wheeler RD, Owens T. Inflammation in the central nervous system and Th17 responses are inhibited by IFN-gamma-Induced IL-18 binding protein. J Immunol. 2010; 185:2458–2466.
Article11. Brakebusch C, Hirsch E, Potocnik A, Fässler R. Genetic analysis of beta1 integrin function: confirmed, new and revised roles for a crucial family of cell adhesion molecules. J Cell Sci. 1997; 110:2895–2904.
Article12. Epler JA, Liu R, Chung H, Ottoson NC, Shimizu Y. Regulation of beta 1 integrin-mediated adhesion by T cell receptor signaling involves ZAP-70 but differs from signaling events that regulate transcriptional activity. J Immunol. 2000; 165:4941–4949.
Article13. Bauer M, Brakebusch C, Coisne C, Sixt M, Wekerle H, Engelhardt B, Fässler R. Beta1 integrins differentially control extravasation of inflammatory cell subsets into the CNS during autoimmunity. Proc Natl Acad Sci USA. 2009; 106:1920–1925.
Article14. Hauzenberger D, Klominek J, Sundqvist KG. Functional specialization of fibronectin-binding beta 1-integrins in T lymphocyte migration. J Immunol. 1994; 153:960–971.15. Baaten BJ, Cooper AM, Swain SL, Bradley LM. Location, location, location: the impact of migratory heterogeneity on T cell function. Front Immunol. 2013; 4:311.
Article16. Loetscher P, Uguccioni M, Bordoli L, Baggiolini M, Moser B, Chizzolini C, Dayer JM. CCR5 is characteristic of Th1 lymphocytes. Nature. 1998; 391:344–345.
Article17. Park HJ, Lee A, Lee JI, Park SH, Ha SJ, Jung KC. Effect of IL-4 on the development and function of memory-like CD8 T cells in the peripheral lymphoid tissues. Immune Netw. 2016; 16:126–133.
Article18. Sorcini D, Bruscoli S, Frammartino T, Cimino M, Mazzon E, Galuppo M, Bramanti P, Al-Banchaabouchi M, Farley D, Ermakova O, et al. Wnt/β-catenin signaling induces integrin α4β1 in T cells and promotes a progressive neuroinflammatory disease in mice. J Immunol. 2017; 199:3031–3041.
Article19. Yang XD, Michie SA, Tisch R, Karin N, Steinman L, McDevitt HO. A predominant role of integrin alpha 4 in the spontaneous development of autoimmune diabetes in nonobese diabetic mice. Proc Natl Acad Sci USA. 1994; 91:12604–12608.
Article20. Jun HK, Lee SH, Lee HR, Choi BK. Integrin α5β1 activates the NLRP3 inflammasome by direct interaction with a bacterial surface protein. Immunity. 2012; 36:755–768.
Article21. Hynes RO. Integrins: a family of cell surface receptors. Cell. 1987; 48:549–554.
Article22. Plow EF, Haas TA, Zhang L, Loftus J, Smith JW. Ligand binding to integrins. J Biol Chem. 2000; 275:21785–21788.
Article23. DeNucci CC, Pagán AJ, Mitchell JS, Shimizu Y. Control of alpha4beta7 integrin expression and CD4 T cell homing by the beta1 integrin subunit. J Immunol. 2010; 184:2458–2467.
Article24. DeNucci CC, Shimizu Y. β1 integrin is critical for the maintenance of antigen-specific CD4 T cells in the bone marrow but not long-term immunological memory. J Immunol. 2011; 186:4019–4026.
Article25. Sixt M, Bauer M, Lämmermann T, Fässler R. Beta1 integrins: zip codes and signaling relay for blood cells. Curr Opin Cell Biol. 2006; 18:482–490.26. Gebhardt T, Wakim LM, Eidsmo L, Reading PC, Heath WR, Carbone FR. Memory T cells in nonlymphoid tissue that provide enhanced local immunity during infection with herpes simplex virus. Nat Immunol. 2009; 10:524–530.
Article27. Chang AC, Salomon DR, Wadsworth S, Hong MJ, Mojcik CF, Otto S, Shevach EM, Coligan JE. Alpha 3 beta 1 and alpha 6 beta 1 integrins mediate laminin/merosin binding and function as costimulatory molecules for human thymocyte proliferation. J Immunol. 1995; 154:500–510.28. Shaw LM, Mercurio AM. Regulation of alpha 6 beta 1 integrin laminin receptor function by the cytoplasmic domain of the alpha 6 subunit. J Cell Biol. 1993; 123:1017–1025.
Article29. Wei J, Shaw LM, Mercurio AM. Integrin signaling in leukocytes: lessons from the alpha6beta1 integrin. J Leukoc Biol. 1997; 61:397–407.
Article30. Krummel MF, Bartumeus F, Gérard A. T cell migration, search strategies and mechanisms. Nat Rev Immunol. 2016; 16:193–201.
Article31. Friedl P, Weigelin B. Interstitial leukocyte migration and immune function. Nat Immunol. 2008; 9:960–969.
Article32. Kang SG, Park J, Cho JY, Ulrich B, Kim CH. Complementary roles of retinoic acid and TGF-β1 in coordinated expression of mucosal integrins by T cells. Mucosal Immunol. 2011; 4:66–82.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Expression of alpha3beta1 Integrin in ECV304 Endothelial Cells and Angiogenesis
- Screening of Integrin Heterodimers Expressed Functionally on the Undifferentiated Spermatogonial Stem Cells in the Outbred ICR Mice
- Integrins Mediating Adhesion and Proliferation of ADP-stimulated Vascular Smooth Muscle Cells
- Inhibition of Transfer Infection of Epstein-Barr Virus to Epithelial Cells by Integrin beta6 siRNA
- Angiostatin Works as Immune Modulatory Molecules via Inhibition of Neutrophil Activation and Migration
