HMG CoA Reductase Inhibitors Inhibit HCV RNA Replication of HCV Genotype 1b but Not 2a
- Affiliations
-
- 1Department of Clinical Laboratory Science, College of Health Sciences, Catholic University of Pusan, Busan, Korea. kschang@cup.ac.kr
- KMID: 2168627
- DOI: http://doi.org/10.4167/jbv.2011.41.2.99
Abstract
- Replication of hepatitis C virus (HCV) is regulated by statin, one of 3-hydroxy-3-methylglutaryl CoA reducatase (HMG CoA reductase) inhibitors that block mevalonate pathway and cholesterol biosyntheis, which has been used usefully for health improvement and disease control in clinic. In order to know which statin can be used to inhibit HCV replication, we examined the effects of HCV genotype 1b replication by 6 kinds of statins with different structure. We treated six statins to HCV genotype 1b replicon cell. Atorvastatin, simvastatin, fluvastatin, mevastatin, and lovastatin inhibited HCV RNA replication and HCV protein expression in HCV genotype 1b replicon cells, though pravastatin did not affect HCV replication. In order to know whether inhibition of HCV replication by statin is depended on HCV genotype, we treated the statins to HCV genotype 2a producing cells, and investigated HCV RNA replication and HCV protein expression. HCV RNA replication and protein expression was not affected in HCV genotype 2a producing cells by treatment of statins and cholesterol inhibitor. These results suggest that HMG-CoA reductase and cholesterol inhibitors might be used depending on HCV genotype. In addition, inhibition of HCV genotype 1b replication by statins has been depended on structure of various statins which should be seriously selected for HCV clinic. In future, we will study on inhibition of another HCV genotype replication by HMG-CoA reductase and cholesterol inhibitors.
MeSH Terms
-
Acyl Coenzyme A
Anticholesteremic Agents
Atorvastatin Calcium
Cholesterol
Fatty Acids, Monounsaturated
Genotype
Hepacivirus
Heptanoic Acids
Hydroxymethylglutaryl CoA Reductases
Hydroxymethylglutaryl-CoA Reductase Inhibitors
Indoles
Lovastatin
Mevalonic Acid
Oxidoreductases
Pravastatin
Pyrroles
Replicon
RNA
Simvastatin
Acyl Coenzyme A
Anticholesteremic Agents
Cholesterol
Fatty Acids, Monounsaturated
Heptanoic Acids
Hydroxymethylglutaryl CoA Reductases
Hydroxymethylglutaryl-CoA Reductase Inhibitors
Indoles
Lovastatin
Mevalonic Acid
Oxidoreductases
Pravastatin
Pyrroles
RNA
Simvastatin
Figure
-
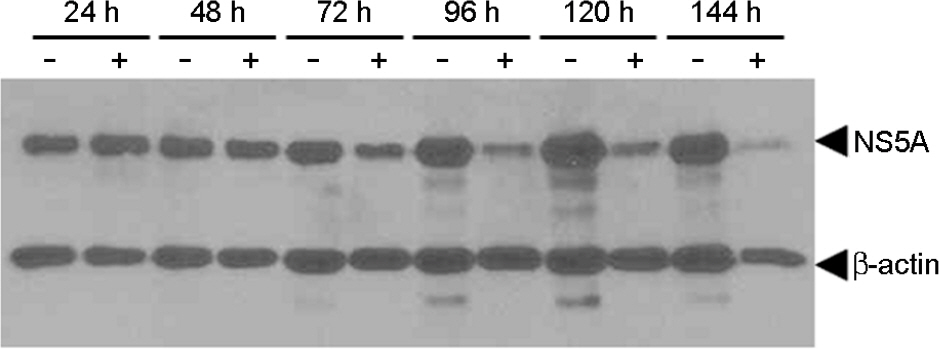
Figure 1. Suppression of HCV genotype 1b RNA replication and HCV protein expression by lovastatin (50 μM), one of HMG-CoA reductase inhibitor, in HCV genotype 1b replicon S1179I #NS3-3 depending on time. Hour is numbered as indicated on the top. Total protein from a clarified lysate (25 μg) was separated by SDS-PAGE and subjected to immunoblot analysis using antibodies directed against NS5A and β-actin. –, not treated; +, treated.
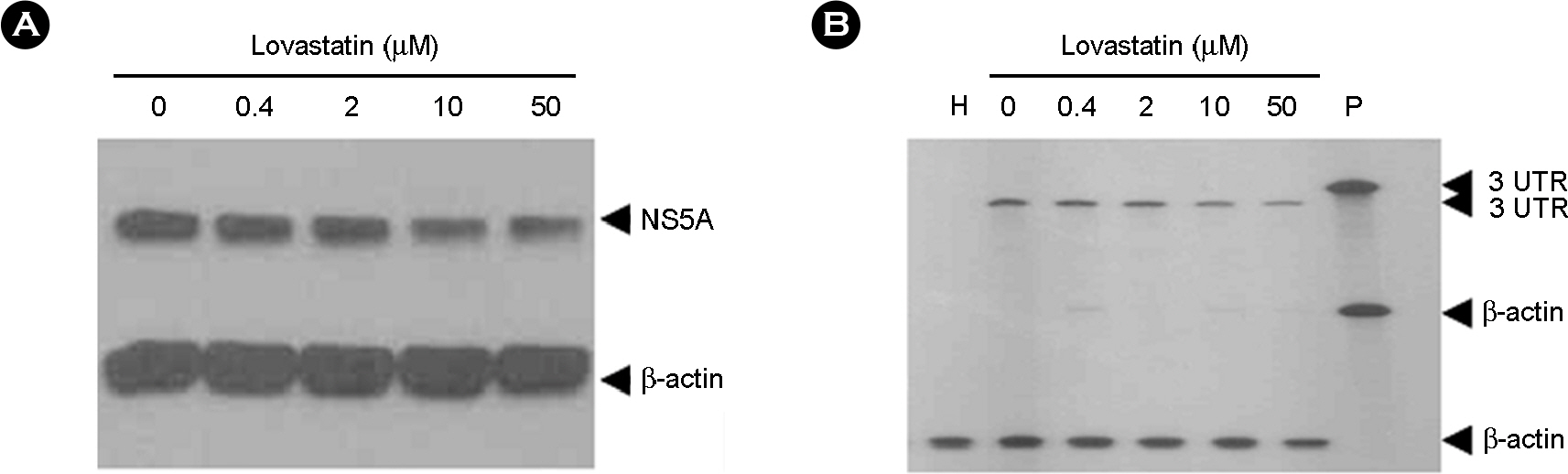
Figure 2. Inhibition of lovastatin to HCV RNA replication and in HCV genotype 1b replicon S1179I #NS3-3 at 72 hrs with increasing concentration. (A) Total protein from a clarified lysate (25 μg) was separated by SDS-PAGE and subjected to immunoblot analysis using antibodies directed against NS5A and β-actin, (B) HCV genotype 1b replicon cell is treated with various concentrations (0, 0.4, 2, 10, and 50 μM) of lovastatin and incubated for 3 days (Lane 2~6, respectively). RNA was extracted using Trizol solution and chloroform. Positive-stranded HCV RNA in each fraction was determined using a negative-sense 3'UTR RNA probe. RNA products were analyzed on a 6% polyacrylamide/7.7 M urea gel and visualized by autoradiography. H, Huh7.5 cell; P, probe.
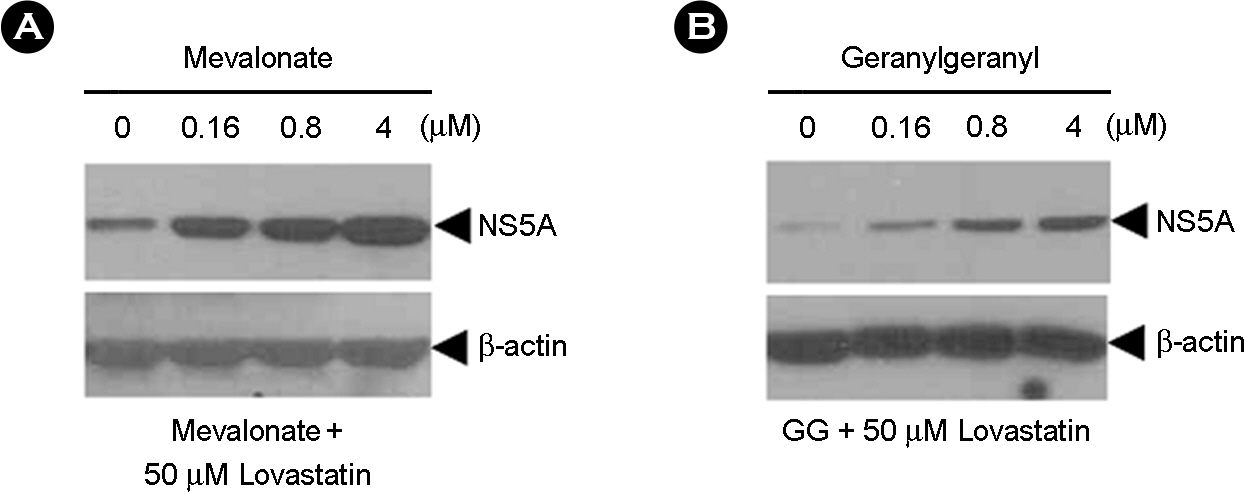
Figure 3. Effects of mevalonate and geranylgeranyl to replication in HCV genotype 1b replicon S1179I #NS3-3 at 72 hrs treated with 50 μM of lovastatin. Total protein from a clarified lysate (25 μg) was separated by SDS-PAGE and subjected to immunoblot analysis using antibodies directed against NS5A and β-actin.

Figure 4. Effects of various statins to HCV replication in HCV genotype 1b replicon S1179I #NS3-3 at 72 hrs by Western blot assay. Total protein from a clarified lysate (25 μg) was separated by SDS-PAGE and subjected to immunoblot analysis using antibodies directed against NS5A and β-actin. ∗cytotoxic effects.
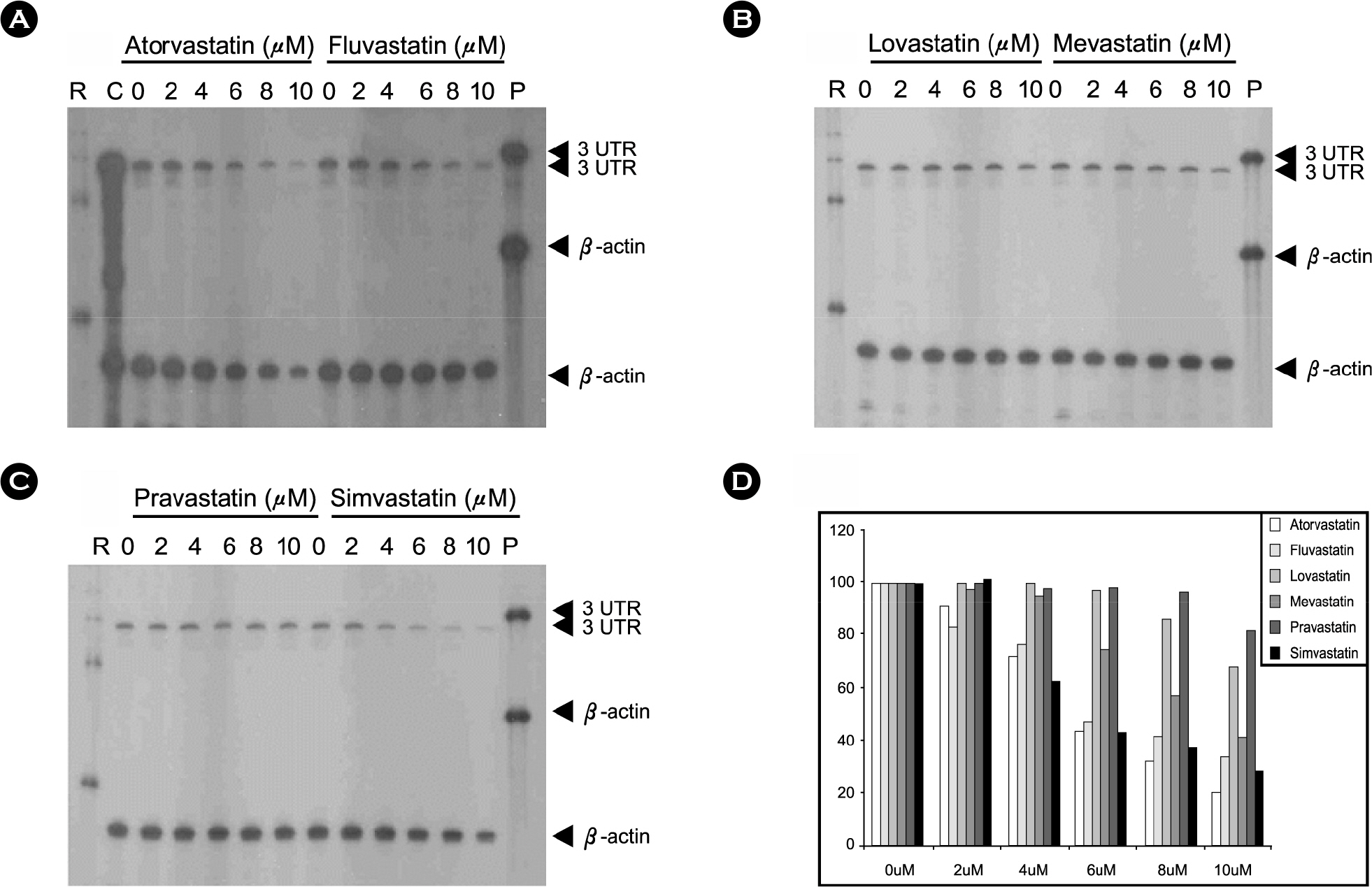
Figure 5. Effects of various statins in HCV genotype 1b replicon S1179I #NS3-3 at 72 hrs by RNase protection assay. HCV genotype 1b replicon cell is treated with various doses (0, 2, 4, 6, 8 and 10 μM) of lovastatin and incubated for 3 days (Lane 2~6, respectively). RNA was extracted using Trizol solution and chloroform. Positive-stranded HCV RNA in each fraction was determined using a negative-sense 3'UTR RNA probe. RNA products were analyzed on a 6% polyacrylamide/7.7 M urea gel and visualized by autoradiography. (D) The intensities of images on gel were quantified using Phosphor imager (Molecular Device). Each experiment was repeated at least three times, and a representative result is shown here. R, RNA marker; P, probe; C, positive control.

Figure 6. Effects of various statins to HCV RNA replication in HCV genotype 2a producing cells at 72 hrs by different dose. (A) Total protein from a clarified lysate (25 μg) was separated by SDS-PAGE and subjected to immunoblot analysis using antibodies directed against NS3 and. β-actin, (B)-(D) HCV genotype 1b replicon cell is treated with different doses (0, 2, 4, 6, 8 and 10 μM) of six statins and incubated for 3 days. RNA was extracted using Trizol solution and chloroform. Positive-stranded HCV RNA in each fraction was determined using a negative-sense 3'UTR RNA probe. RNA products were analyzed on a 6% polyacrylamide/7.7 M urea gel and visualized by autoradiography. R, RNA marker; P, probe; NT, not treated (negative control).
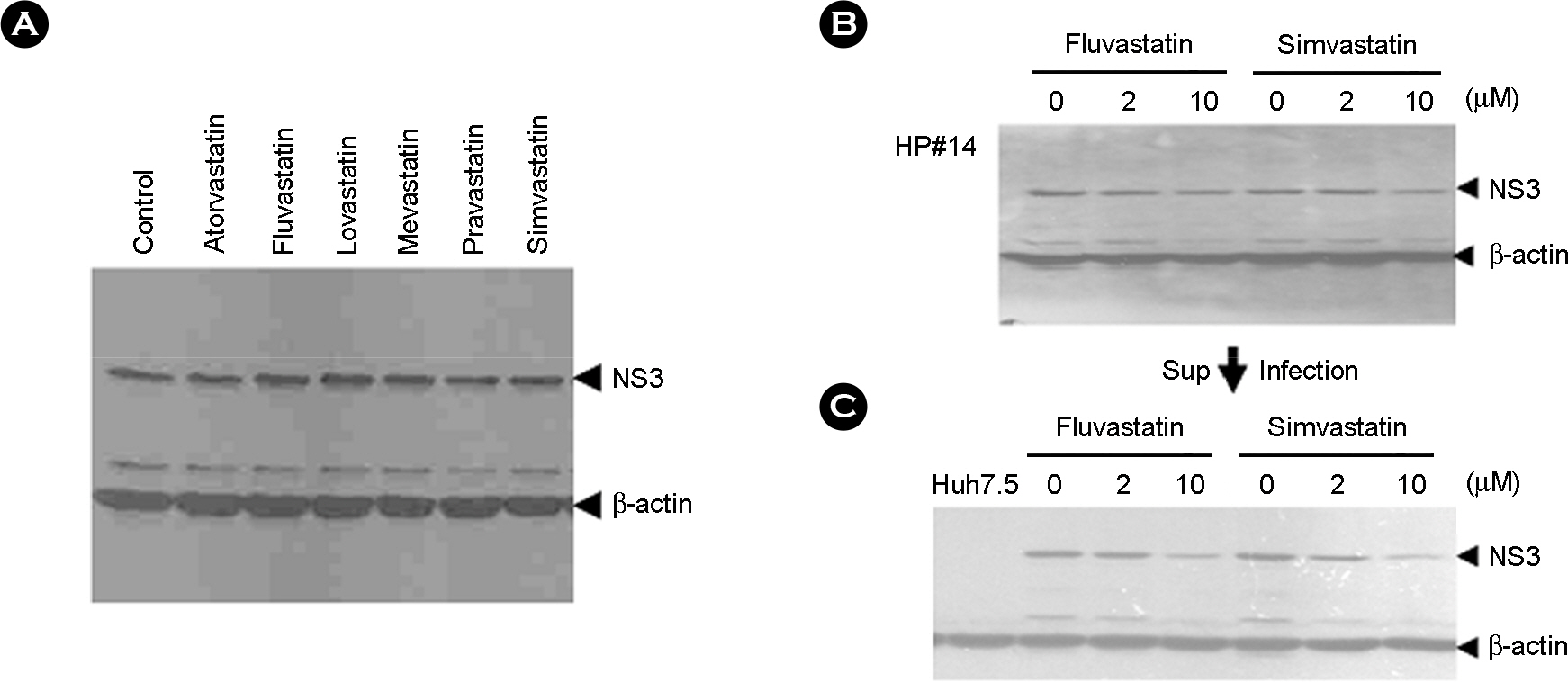
Figure 7. Effects of various statins to HCV protein expression in HCV genotype 2a producing cell lines at 72 hrs by different concentration. Total protein from a clarified lysate (25 μg) was separated by SDS-PAGE and subjected to immunoblot analysis using antibodies directed against NS3 and β-actin. (A) NS3 protein expression in HCV genetype producing cell treated with 50 μM of six statins, (B) Effects to HCV replication in HCV producing cell treated with 2 and 10 μM of fluvastatin and simvastatin, (C) Effects to HCV infection in Huh7.5 infected with the supernatant from HCV producing cell treated with fluvastatin and simvastatin.

Figure 8. Effects of HCV replication in HCV genotype 1b replicon S1179I #NS3-3 (A) and HCV genotype 2a replicon (B) treated with different concentration of MβCD at 72 hrs. Total protein from a clarified lysate (25 μg) was separated by SDS-PAGE and subjected to immunoblot analysis using antibodies directed against NS5A (A), NS3 (B) and β-actin.

Figure 9. Effect of MβCD to HCV RNA replication in HCV genotype 2a producing cell lines at 72 hrs by different concentration. Total protein from a clarified lysate (25 μg) was separated by SDS-PAGE and subjected to immunoblot analysis using antibodies directed against NS3 and β-actin. (A) Effects to HCV replication in HCV producing cell treated with MβCD. (B). Effects to HCV infection in Huh7.5 infected with the sup. AI, treatment after infection; BI, treatment before infectio; NI, not infected; HS, supernatant from Huh7.5 cell. ∗cytotoxic effects.
Reference
-
1). Alter MJ. Epidemiology of hepatitis C. Hepatology. 1997. 26:62S–5S.
Article2). McHutchison JG., Fried MW. Current therapy for hepatitis C: pegylated interferon and ribavirin. Clin Liver Dis. 2003. 7:149–61.
Article3). Lindebach BD., Rice CM. Fields virology. Knipe DM, Howley PM, editors. editors.The hepatitis C viruses. 4th ed.Philadelphia: Lippincott Williams & Wilkins;2002. p. 602–13.4). Kapadia SB., Chisari FV. Hepatitis C virus RNA replication is regulated by host geranylgeranylation and fatty acids. Proc Natl Acad Sci U S A. 2005. 102:2561–6.
Article5). Ye J., Wang C., Sumpter R Jr., Brown MS., Goldstein JL., Gale M Jr. Disruption of hepatitis C virus RNA replication through inhibition of host protein geranyl-geranylation. Proc Natl Acad Sci U S A. 2003. 100:15865–70.
Article6). Ikeda M., Abe K., Yamada M., Dansako H., Naka K., Kato N. Different anti-HCV profiles of statins and their potential for combination therapy with interferon. Hepatology. 2006. 44:117–25.
Article7). Ikeda M., Kato N. Life style-related diseases of the digestive system: cell culture system for the screening of anti-hepatitis C virus (HCV) reagents: suppression of HCV replication by statins and synergistic action with interferon. J Pharmacol Sci. 2007. 105:145–50.
Article8). Verma SP. HIV: a raft-targeting approach for prevention and therapy using plant-derived compounds (review). Curr Drug Targets. 2009. 10:51–9.
Article9). Wang W., Fu YJ., Zu YG., Wu N., Reichling J., Efferth T. Lipid rafts play an important role in the vesicular stomatitis virus life cycle. Arch Virol. 2009. 154:595–600.
Article10). Pollock S., Nichita NB., Böhmer A., Radulescu C., Dwek RA., Zitzmann N. Polyunsaturated liposomes are antiviral against hepatitis B and C viruses and HIV by decreasing cholesterol levels in infected cells. Proc Natl Acad Sci U S A. 2010. 107:17176–81.
Article11). Cianciola NL., Carlin CR. Adenovirus RID-alpha activates an autonomous cholesterol regulatory mechanism that rescues defects linked to Niemann-Pick disease type C. J Cell Biol. 2009. 187:537–52.12). Forde KA., Law C., O'Flynn R., Kaplan DE. Do statins reduce hepatitis C RNA titers during routine clinical use? World J Gastroenterol. 2009. 15:5020–7.
Article13). O'Leary JG., Chan JL., McMahon CM., Chung RT. Atorvastatin does not exhibit antiviral activity against HCV at conventional doses: a pilot clinical trial. Hepatology. 2007. 45:895–8.14). Luo G., Xin S., Cai Z. Role of the 5′-proximal stem-loop structure of the 5′ untranslated region in replication and translation of hepatitis C virus RNA. J Virol. 2003. 77:3312–8.
Article15). Cai Z., Zhang C., Chang KS., Jiang J., Ahn BC., Wakita T, et al. Robust production of infectious hepatitis C virus (HCV) from stably HCV cDNA-transfected human hepatoma cells. J Virol. 2005. 79:13963–73.
Article16). Chang KS., Luo G. The polypyrimidine tract-binding protein (PTB) is required for efficient replication of hepatitis C virus (HCV) RNA. Virus Res. 2006. 115:1–8.
Article17). He Z., Zhuang H., Wang X., Song S., Dong Q., Yan J, et al. Retrospective analysis of non-A-E hepatitis: possible role of hepatitis B and C virus infection. J Med Virol. 2003. 69:59–65.
Article18). Chang KS., Jiang J., Cai Z., Luo G. Human apolipoprotein E is required for infectivity and production of hepatitis C virus in cell culture. J Virol. 2007. 81:13783–93.
Article19). Khorashadi S., Hasson NK., Cheung RC. Incidence of statin hepatotoxicity in patients with hepatitis C. Clin Gastroenterol Hepatol. 2006. 4:902–7.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Hepatitis C Virus Genotypes in Korean Hemophiliacs
- Distribution of Hepatitis C Virus Genotypes among Intravenous Drug Users
- Genotypes of Hepatitis C Virus and Short Term Efficacy of alpha-nterferon Therapy in Patients with HCV Infection in Taegu
- Apolipoprotein E Genotype and Expression Correlated with Hepatitis C Virus Genotype and Infection
- Distribution of hepatitis C virus genotypes determined by line probe assay in Korean patients with chronic HCV infection
