Recombinant TAT-CD137 Ligand Cytoplasmic Domain Fusion Protein Induces the Production of IL-6 and TNF-alpha in Peritoneal Macrophages
- Affiliations
-
- 1School of Biological Sciences, Ulsan University Hospital, University of Ulsan School of Medicine, Ulsan 680-749, Korea. bkwon@mail.ulsan.ac.kr
- 2Department of Surgery, Ulsan University Hospital, University of Ulsan School of Medicine, Ulsan 680-749, Korea.
- 3Biomedical Research Center, Ulsan University Hospital, University of Ulsan School of Medicine, Ulsan 680-749, Korea.
- KMID: 1449813
- DOI: http://doi.org/10.4110/in.2011.11.4.216
Abstract
- BACKGROUND
The ligand for CD137 (CD137L; also called 4-1BBL) is mainly expressed on activated APCs such as dendritic cells, B cells and macrophages. Even though CD137L functions as a trigger of the CD137 signaling pathway for T cell activation and expansion, engagement of CD137L can deliver a signal leading to the production of proinflammatory cytokines in macrophages.
METHODS
We generated cell-permeable TAT-CD137L cytoplasmic domain fusion protein (TAT-CD137Lct) and examined its ability to initiate the CD137L reverse signaling pathway.
RESULTS
Treatment of TAT-CD137Lct induced the production of high levels of IL-6 and TNF-alpha mRNAs and proteins in peritoneal macrophages. TAT-CD137Lct increased phosphorylation of Erk, p38 MAPK and Jnk, and activated transcription factors C/EBP and CREB. However, TAT-CD137Lct did not visibly affect the degradation of the inhibitor of NF-kB (IkBalpha). We further demonstrated that JNK activation was required for TAT-CD137Lct-induced production of TNF-alpha, while activation of Erk and p38 MAPK were involved in IL-6 and TNF-alpha production.
CONCLUSION
Our results suggest that TAT-CD137Lct is an effective activator for the CD137L reverse signaling pathway.
Keyword
MeSH Terms
-
4-1BB Ligand
B-Lymphocytes
Cytokines
Cytoplasm
Dendritic Cells
Interleukin-6
Macrophages
Macrophages, Peritoneal
NF-kappa B
p38 Mitogen-Activated Protein Kinases
Phosphorylation
Proteins
RNA, Messenger
Transcription Factors
Tumor Necrosis Factor-alpha
4-1BB Ligand
Cytokines
Interleukin-6
NF-kappa B
Proteins
RNA, Messenger
Transcription Factors
Tumor Necrosis Factor-alpha
p38 Mitogen-Activated Protein Kinases
Figure
-
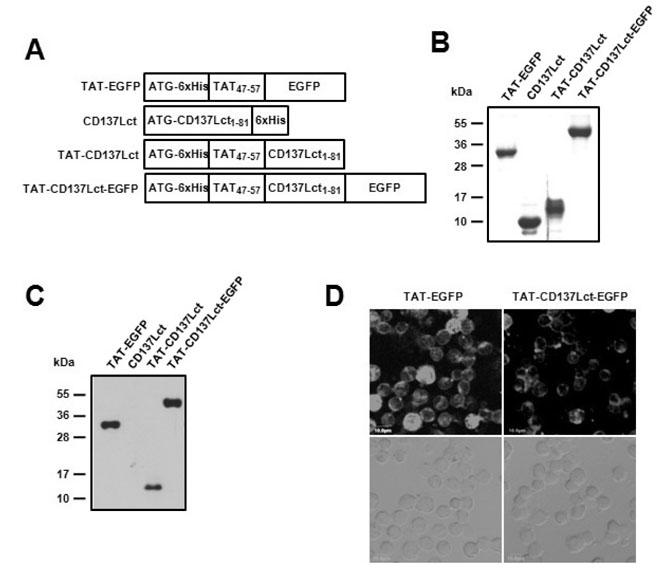
Figure 1 Production and transduction of TAT fusion proteins. (A) Plasmid constructs of the TAT fusion proteins. (B) Purified proteins were separated on 13% SDS-PAGE and then stained with Commasie blue. (C, D) Transduction activity of purified proteins was analyzed using immunoblot (C) and confocal microscope (D). Peritoneal macrophages (1×106) were incubated with 10 µM of TAT-EGFP, CD137Lct, TAT-CD137Lct, and TAT-CD137Lct-EGFP for 30 min. Transduced proteins were detected using anti-his Abs (C). Intracellular localization of TAT-EGFP and TAT-CD137Lct-EGFP were observed using confocal microscopy (D). Scale bar is 10 µm.
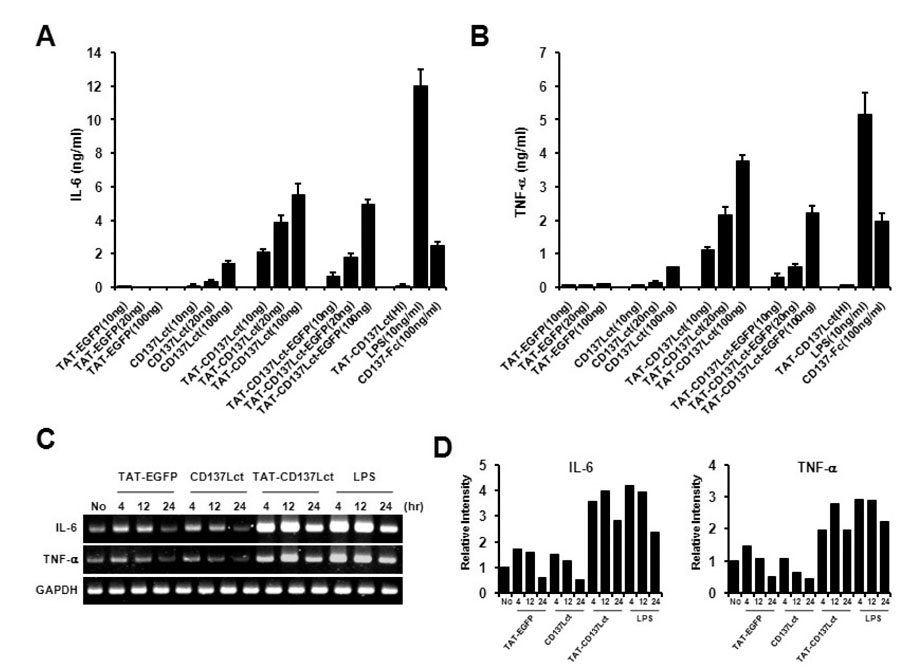
Figure 2 TAT-CD137Lct induces IL-6 and TNF production in peritoneal macrophages. (A, B) Peritoneal macrophages (1×105) were treated with indicated concentrations of TAT- EGFP, CD137Lct, TAT-CD137Lct, and TAT-CD137Lct-EGFP and cultured for 4 h. Culture medium was collected and levels of IL-6 (A) and TNF-α (B) were measured using a CBA kit. Heat-inactivated (HI) TAT-CD137Lct was used as a negative control and LPS (10 ng/ml) and CD137-Fc (100 ng/ml) as a positive control. Data are the mean±SD of triplicates. (C) Total RNA was extracted from peritoneal macrophages (1×106) that were treated with 20 ng of protein. Total RNA was subjected to cDNA synthesis. The TNF-α and IL-6 transcripts were amplified using specific primer sets and the PCR products were analyzed using 1.5% agarose gel electrophoresis. GAPDH mRNA expression was used as an internal control. (D) Density of amplified PCR product bands was analyzed using Image J program and normalized against the density of the GAPDH band.
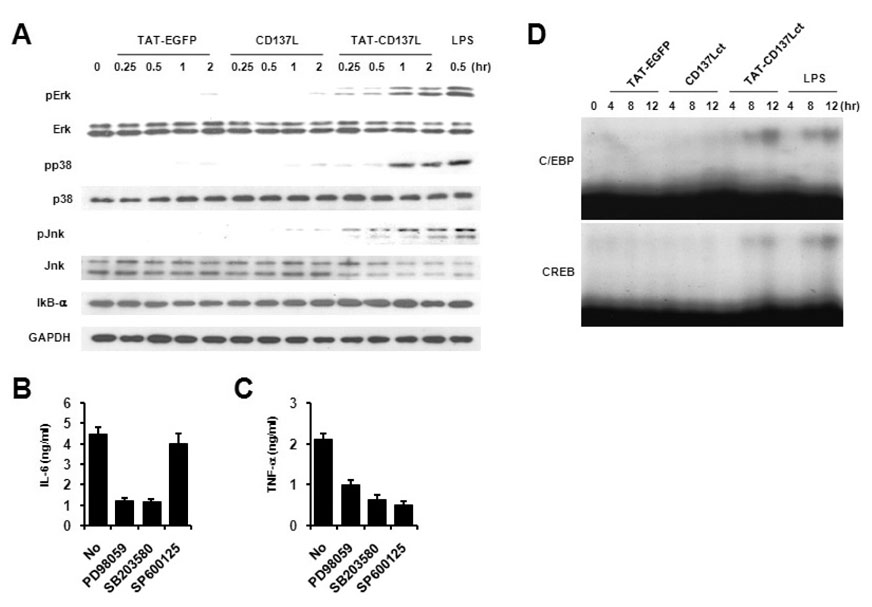
Figure 3 TAT-CD137Lct induced phosphorylation of Erk, p38, and Jnk and activaton of C/EBP and CREB. (A) Peritoneal macrophages (1×106) were treated with 50 ng of indicated proteins and harvested at 0, 15, 30, 60, and 120 min after their treatment. Lysates (10 µg) were used for immunoblot. LPS (50 ng/ml) is used as a positive control. p-, phosphorylated. (B, C) Peritoneal macrophages (1×105) were treated with TAT-CD137Lct (20 ng) in the presence of PD98059, SB203580, or SP600125 (10 µM each) and cultured for 4 h. Levels of IL-6 (B) and TNF (C) in culture medium were measured using a CBA kit. Data are the mean±SD of triplicates. (D) EMSA for nuclear extracts from peritoneal macrophages stimulated with 50 ng of indicated proteins was performed with radiolabeled C/EBP and CREB probes. LPS (50 ng/ml) is used as a positive control.

Figure 4 An additive effect of TAT-CD137Lct and LPS. Peritoneal macrophages (1×105) were treated with indicated concentrations of TAT-CD137Lct and LPS and cultured for 4 h. Culture medium was collected and levels of IL-6 (A) and TNF-α (B) was measured using a CBA kit.
Cited by 1 articles
-
Regulation of Inflammation by Bidirectional Signaling through CD137 and Its Ligand
Byungsuk Kwon
Immune Netw. 2012;12(5):176-180. doi: 10.4110/in.2012.12.5.176.
Reference
-
1. Eissner G, Kolch W, Scheurich P. Ligands working as receptors: reverse signaling by members of the TNF superfamily enhance the plasticity of the immune system. Cytokine Growth Factor Rev. 2004. 15:353–366.
Article2. Schwarz H. Biological activities of reverse signal transduction through CD137 ligand. J Leukoc Biol. 2005. 77:281–286.
Article3. Shao Z, Schwarz H. CD137 ligand, a member of the tumor necrosis factor family, regulates immune responses via reverse signal transduction. J Leukoc Biol. 2011. 89:21–29.
Article4. Langstein J, Michel J, Fritsche J, Kreutz M, Andreesen R, Schwarz H. CD137 (ILA/4-1BB), a member of the TNF receptor family, induces monocyte activation via bidirectional signaling. J Immunol. 1998. 160:2488–2494.5. Langstein J, Michel J, Schwarz H. CD137 induces proliferation and endomitosis in monocytes. Blood. 1999. 94:3161–3168.
Article6. Langstein J, Schwarz H. Identification of CD137 as a potent monocyte survival factor. J Leukoc Biol. 1999. 65:829–833.
Article7. Langstein J, Becke FM, Söllner L, Krause G, Brockhoff G, Kreutz M, Andreesen R, Schwarz H. Comparative analysis of CD137 and LPS effects on monocyte activation, survival, and proliferation. Biochem Biophys Res Commun. 2000. 273:117–122.
Article8. Ju SW, Ju SG, Wang FM, Gu ZJ, Qiu YH, Yu GH, Ma HB, Zhang XG. A functional anti-human 4-1BB ligand monoclonal antibody that enhances proliferation of monocytes by reverse signaling of 4-1BBL. Hybrid Hybridomics. 2003. 22:333–338.
Article9. Laderach D, Wesa A, Galy A. 4-1BB-ligand is regulated on human dendritic cells and induces the production of IL-12. Cell Immunol. 2003. 226:37–44.
Article10. Lippert U, Zachmann K, Ferrari DM, Schwarz H, Brunner E, Mahbub-Ul Latif AH, Neumann C, Soruri A. CD137 ligand reverse signaling has multiple functions in human dendritic cells during an adaptive immune response. Eur J Immunol. 2008. 38:1024–1032.
Article11. Schwarz H, Blanco FJ, von Kempis J, Valbracht J, Lotz M. ILA, a member of the human nerve growth factor/tumor necrosis factor receptor family, regulates T-lymphocyte proliferation and survival. Blood. 1996. 87:2839–2845.
Article12. Kwajah M M S, Mustafa N, Holme AL, Pervaiz S, Schwarz H. Biphasic activity of CD137 ligand-stimulated monocytes on T cell apoptosis and proliferation. J Leukoc Biol. 2011. 89:707–720.
Article13. Pauly S, Broll K, Wittmann M, Giegerich G, Schwarz H. CD137 is expressed by follicular dendritic cells and costimulates B lymphocyte activation in germinal centers. J Leukoc Biol. 2002. 72:35–42.14. Pollok KE, Kim YJ, Hurtado J, Zhou Z, Kim KK, Kwon BS. 4-1BB T-cell antigen binds to mature B cells and macrophages, and costimulates anti-mu-primed splenic B cells. Eur J Immunol. 1994. 24:367–374.
Article15. Jeon HJ, Choi JH, Jung IH, Park JG, Lee MR, Lee MN, Kim B, Yoo JY, Jeong SJ, Kim DY, Park JE, Park HY, Kwack K, Choi BK, Kwon BS, Oh GT. CD137 (4-1BB) deficiency reduces atherosclerosis in hyperlipidemic mice. Circulation. 2010. 121:1124–1133.
Article16. Salih HR, Kosowski SG, Haluska VF, Starling GC, Loo DT, Lee F, Aruffo AA, Trail PA, Kiener PA. Constitutive expression of functional 4-1BB (CD137) ligand on carcinoma cells. J Immunol. 2000. 165:2903–2910.
Article17. Gullo C, Koh LK, Pang WL, Ho KT, Tan SH, Schwarz H. Inhibition of proliferation and induction of apoptosis in multiple myeloma cell lines by CD137 ligand signaling. PLoS One. 2010. 5:e10845.
Article18. Drenkard D, Becke FM, Langstein J, Spruss T, Kunz-Schughart LA, Tan TE, Lim YC, Schwarz H. CD137 is expressed on blood vessel walls at sites of inflammation and enhances monocyte migratory activity. FASEB J. 2007. 21:456–463.
Article19. Kang YJ, Kim SO, Shimada S, Otsuka M, Seit-Nebi A, Kwon BS, Watts TH, Han J. Cell surface 4-1BBL mediates sequential signaling pathways 'downstream' of TLR and is required for sustained TNF production in macrophages. Nat Immunol. 2007. 8:601–609.
Article20. Lee SW, Park Y, So T, Kwon BS, Cheroutre H, Mittler RS, Croft M. Identification of regulatory functions for 4-1BB and 4-1BBL in myelopoiesis and the development of dendritic cells. Nat Immunol. 2008. 9:917–926.
Article21. Jiang D, Chen Y, Schwarz H. CD137 induces proliferation of murine hematopoietic progenitor cells and differentiation to macrophages. J Immunol. 2008. 181:3923–3932.
Article22. Jiang D, Yue PS, Drenkard D, Schwarz H. Induction of proliferation and monocytic differentiation of human CD34+ cells by CD137 ligand signaling. Stem Cells. 2008. 26:2372–2381.
Article23. Asoh S, Ohta S. PTD-mediated delivery of anti-cell death proteins/peptides and therapeutic enzymes. Adv Drug Deliv Rev. 2008. 60:499–516.
Article24. Choi JM, Ahn MH, Chae WJ, Jung YG, Park JC, Song HM, Kim YE, Shin JA, Park CS, Park JW, Park TK, Lee JH, Seo BF, Kim KD, Kim ES, Lee DH, Lee SK, Lee SK. Intranasal delivery of the cytoplasmic domain of CTLA-4 using a novel protein transduction domain prevents allergic inflammation. Nat Med. 2006. 12:574–579.
Article25. Choi JM, Kim SH, Shin JH, Gibson T, Yoon BS, Lee DH, Lee SK, Bothwell AL, Lim JS, Lee SK. Transduction of the cytoplasmic domain of CTLA-4 inhibits TcR-specific activation signals and prevents collagen-induced arthritis. Proc Natl Acad Sci U S A. 2008. 105:19875–19880.
Article26. Kwon B. CD137-CD137 Ligand Interactions in Inflammation. Immune Netw. 2009. 9:84–89.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Effects of cytokines in the activation of peritoneal macrophages from mice infected with Toxoplasma gondii
- Regulation of Inflammation by Bidirectional Signaling through CD137 and Its Ligand
- The Production and Correlation of Silica Induced Proinflammatory Cytokines and TGF-beta from Monocytes of Balb/C Mice
- Metformin Down-regulates TNF-alpha Secretion via Suppression of Scavenger Receptors in Macrophages
- The Role of Intracellular Receptor NODs for Cytokine Production by Macrophages Infected with Mycobacterium leprae
