CD40 Co-stimulation Inhibits Sustained BCR-induced Ca2+ Signaling in Response to Long-term Antigenic Stimulation of Immature B Cells
- Affiliations
-
- 1Department of Physiology, SBRI, Sungkyunkwan University School of Medicine, Suwon 440-746, Korea. tongmkang@skku.edu
- 2Department of Molecular Cell Biology, SBRI, Sungkyunkwan University School of Medicine, Suwon 440-746, Korea.
- 3Department of Physiology, Seoul National University College of Medicine, Seoul 110-799, Korea.
- KMID: 1448922
- DOI: http://doi.org/10.4196/kjpp.2011.15.3.179
Abstract
- Regulation of B cell receptor (BCR)-induced Ca2+ signaling by CD40 co-stimulation was compared in long-term BCR-stimulated immature (WEHI-231) and mature (Bal-17) B cells. In response to long-term pre-stimulation of immature WEHI-231 cells to alpha-IgM antibody (0.5~48 hr), the initial transient decrease in BCR-induced [Ca2+]i was followed by spontaneous recovery to control level within 24 hr. The recovery of Ca2+ signaling in WEHI-231 cells was not due to restoration of internalized receptor but instead to an increase in the levels of PLCgamma2 and IP3R-3. CD40 co-stimulation of WEHI-231 cells prevented BCR-induced cell cycle arrest and apoptosis, and it strongly inhibited the recovery of BCR-induced Ca2+ signaling. CD40 co-stimulation also enhanced BCR internalization and reduced expression of PLCgamma2 and IP3R-3. Pre-treatment of WEHI-231 cells with the antioxidant N-acetyl-L-cysteine (NAC) strongly inhibited CD40-mediated prevention of the recovery of Ca2+ signaling. In contrast to immature WEHI-231 cells, identical long-term alpha-IgM pre-stimulation of mature Bal-17 cells abolished the increase in BCR-induced [Ca2+]i, regardless of CD40 co-stimulation. These results suggest that CD40-mediated signaling prevents antigen-induced cell cycle arrest and apoptosis of immature B cells through inhibition of sustained BCR-induced Ca2+ signaling.
Keyword
MeSH Terms
Figure
-
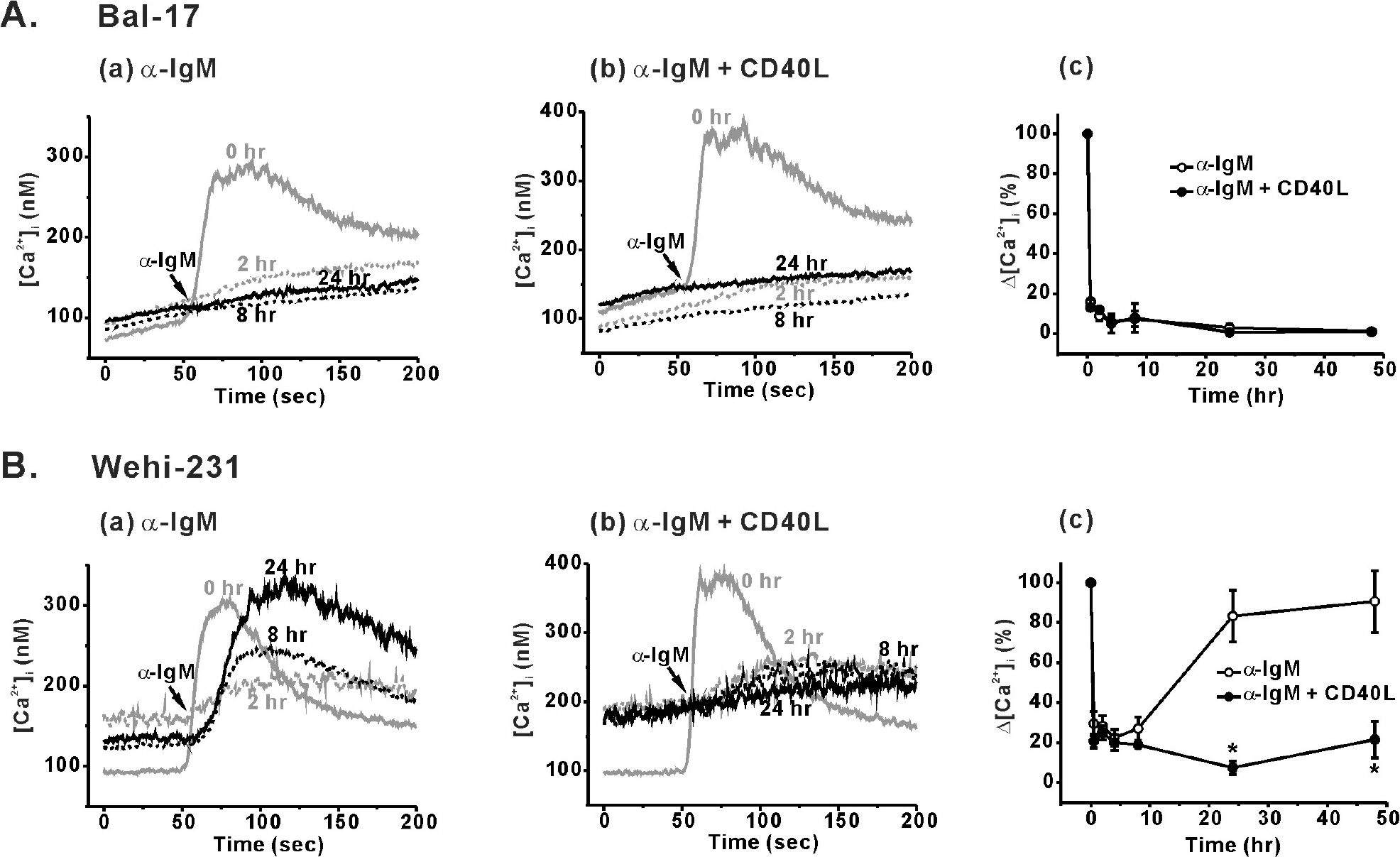
Fig. 1. Differential BCR-induced Ca2+ responses and its modulation by CD40 co-stimulation in Bal-17 and WEHI-231 cells. B cells were pretreated with 5 μg/ml of α-IgM (Aa, Ba) or α-IgM + CD40L (0.5 μg/ml) (Ab, Bb) for 0.5, 2, 4, 8, 24, and 48 hr. After harvesting the cells, the pre-treatment agents were removed, and α-IgM (5 μg/ml)-triggered [Ca2+]i increases were measured in 1.5 mM [Ca2+]o containing normal Tyrode solution. Representative [Ca2+]i traces obtained from each B cell type are drawn, and the duration of α-IgM pre-stimulation (0, 2, 8, and 24 hr) are indicated inside the figure. (Ac, Bc) Δ[Ca2+]i values (difference between the resting and peak [Ca2+]i levels) were calculated and normalized against Δ[Ca2+]i values of un-stimulated control cells. Changes in Δ[Ca2+]i values are plotted against the duration of pre-stimulation. All experiments were repeated at least three times, and the detailed values are described in the text. ∗p<0.05.

Fig. 2. Contribution of cell membrane potential, amounts of intracellular Ca2+ release and store-operated Ca2+ entry (SOC) on sustained BCR-induced Ca2+ responses in immature WEHI-231 cells. WEHI-231 cells were pretreated with 5 μg/ ml of α-IgM or α-IgM+CD40L (0.5 μg/ml) for 24 hr. After harvesting the cells, the pre-treatment agents were removed, and α-IgM (5 μg/ml)-triggered [Ca2+]i increases were measured in 130 mM [KCl]o and 1.5 mM [Ca2+]o containing solution (A) or extracellular Ca2+-free Tyrode solution (B). The amounts of store-operated Ca2+ entry in response to the stimulation of the cells with α-IgM (B) or thapsigargin (C) were measured by the re-addition of 1.5 mM [Ca2+]o after depleting the stores in Ca2+-free Tyrode solution. (D) Δ[Ca2+]i values (difference between the resting and peak [Ca2+]i levels) from (A), (B) and (C) were calculated and plotted. Mean values±S.E.M. from at least 3 repeats were shown. #p<0.05.
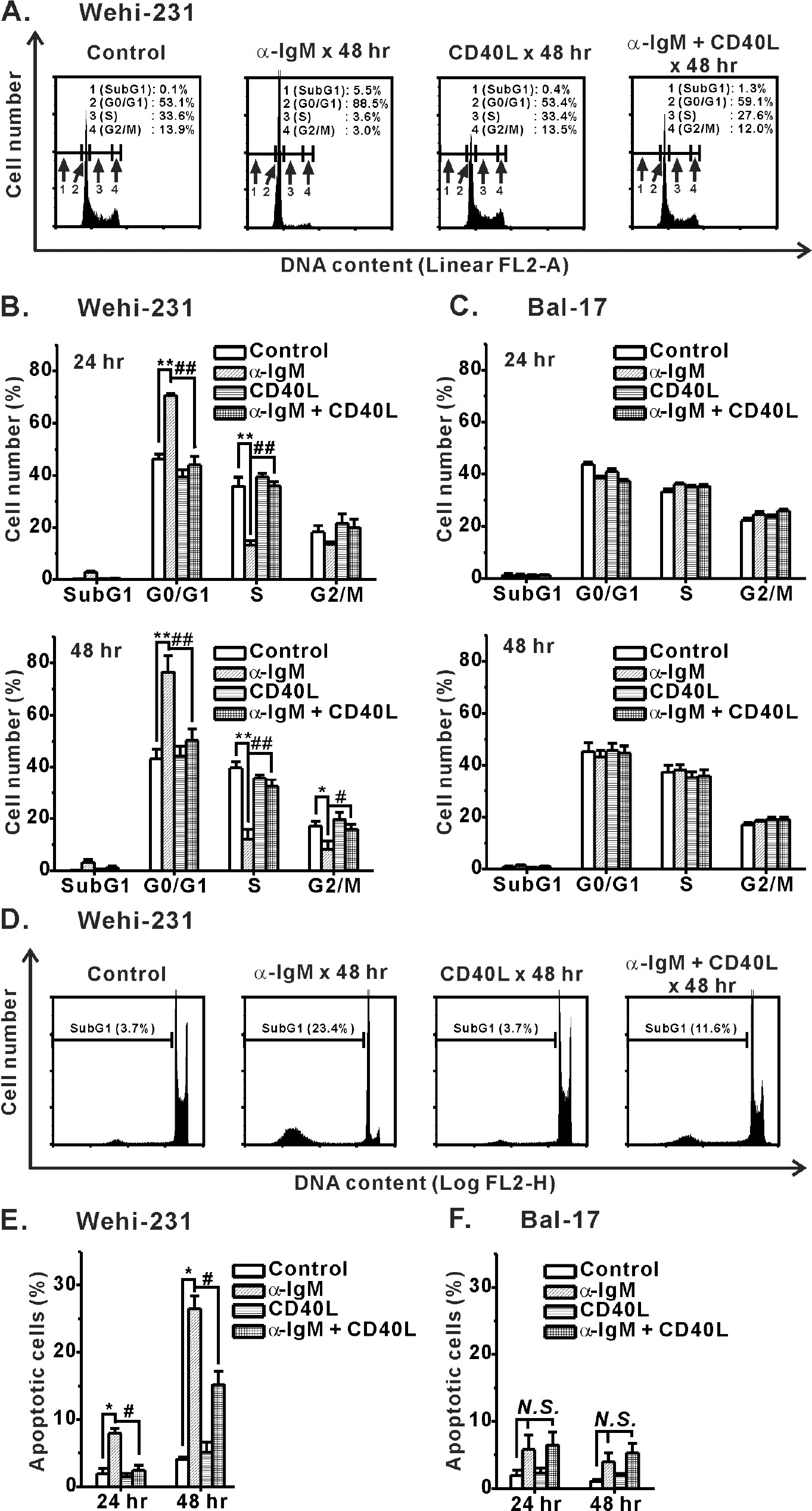
Fig. 3. Inhibition of α-IgM-induced cell cycle arrest and apoptosis of immature WEHI-231 cells by CD40 co-stimulation. WEHI-231 and Bal-17 cells were stimulated with α-IgM or α-IgM+CD40L for 24 and 48 hr, after which FACS analysis was performed to quantify the degree of cell cycle arrest (A∼C) and apoptosis (D∼F). Cell cycle and apoptosis were analyzed on linear FL2-area (FL2-A, total cell PI fluorescence) and logarithmic FL2-height (FL2-H, maximum PI fluorescence emission) scales, respectively. All experiments were repeated three times. Mean±S.E.M. ∗,#p<0.05, ∗∗,##p<0.01, N.S., no significance.
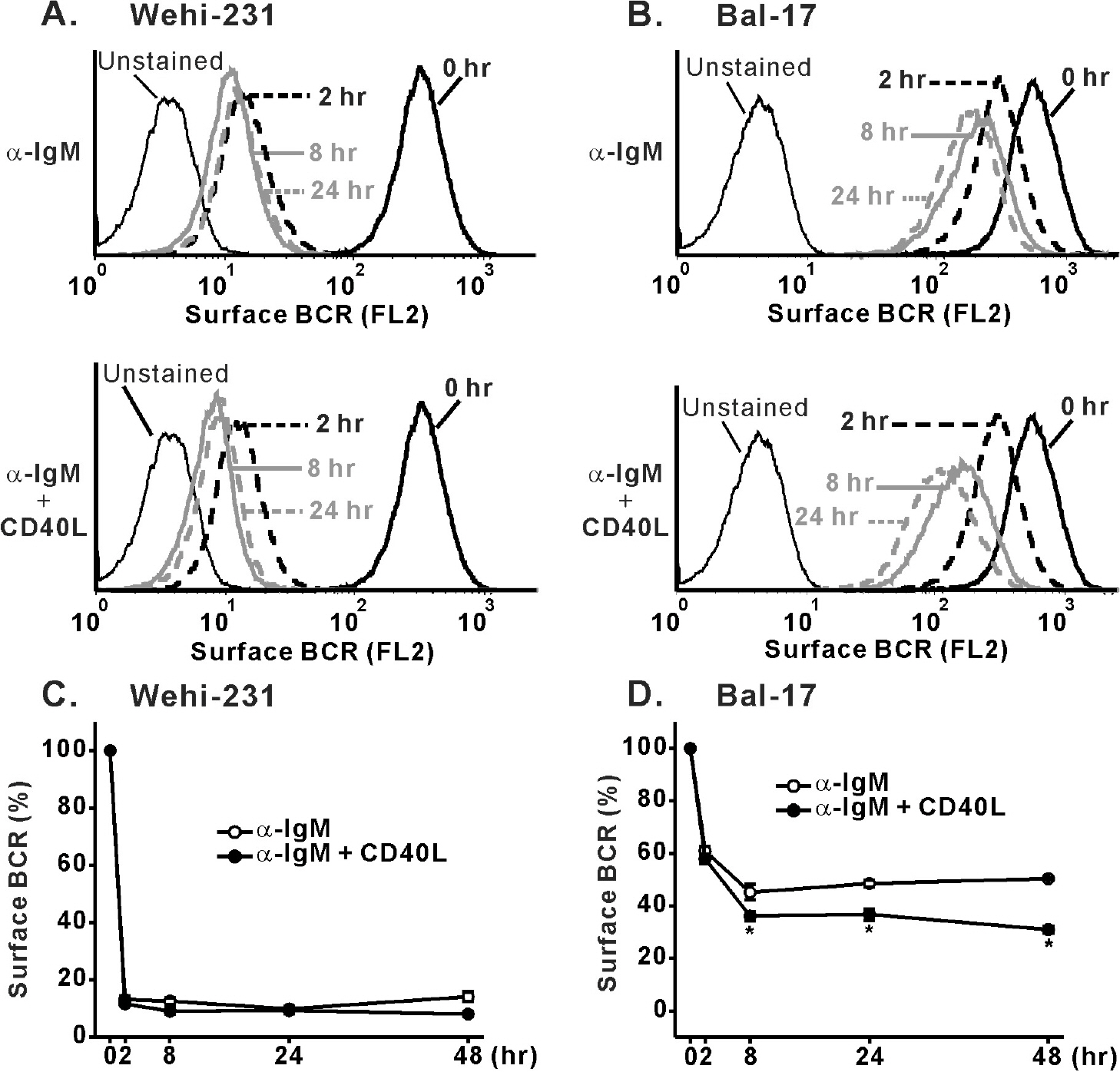
Fig. 4. Effects of CD40 co-stimulation on internalization of surface BCR triggered by α-IgM pre-stimulation. WEHI-231 (A) and Bal-17 (B) cells were stimulated with α-IgM alone or α-IgM + CD40L for the indicated times, followed by surface BCR measurement indicated by PE fluorescence emission (FL2-H) in FACS machine. Effects of CD40 co-stimulation on α-IgM-triggered internalization of BCR were quantified and normalized (% surface BCR) to that of un-treated cells and then plotted against the indicated times of stimulation in WEHI-231 (C) and Bal-17 (D) cells. Mean±S.E.M. values of three repeats are plotted. ∗p<0.05.
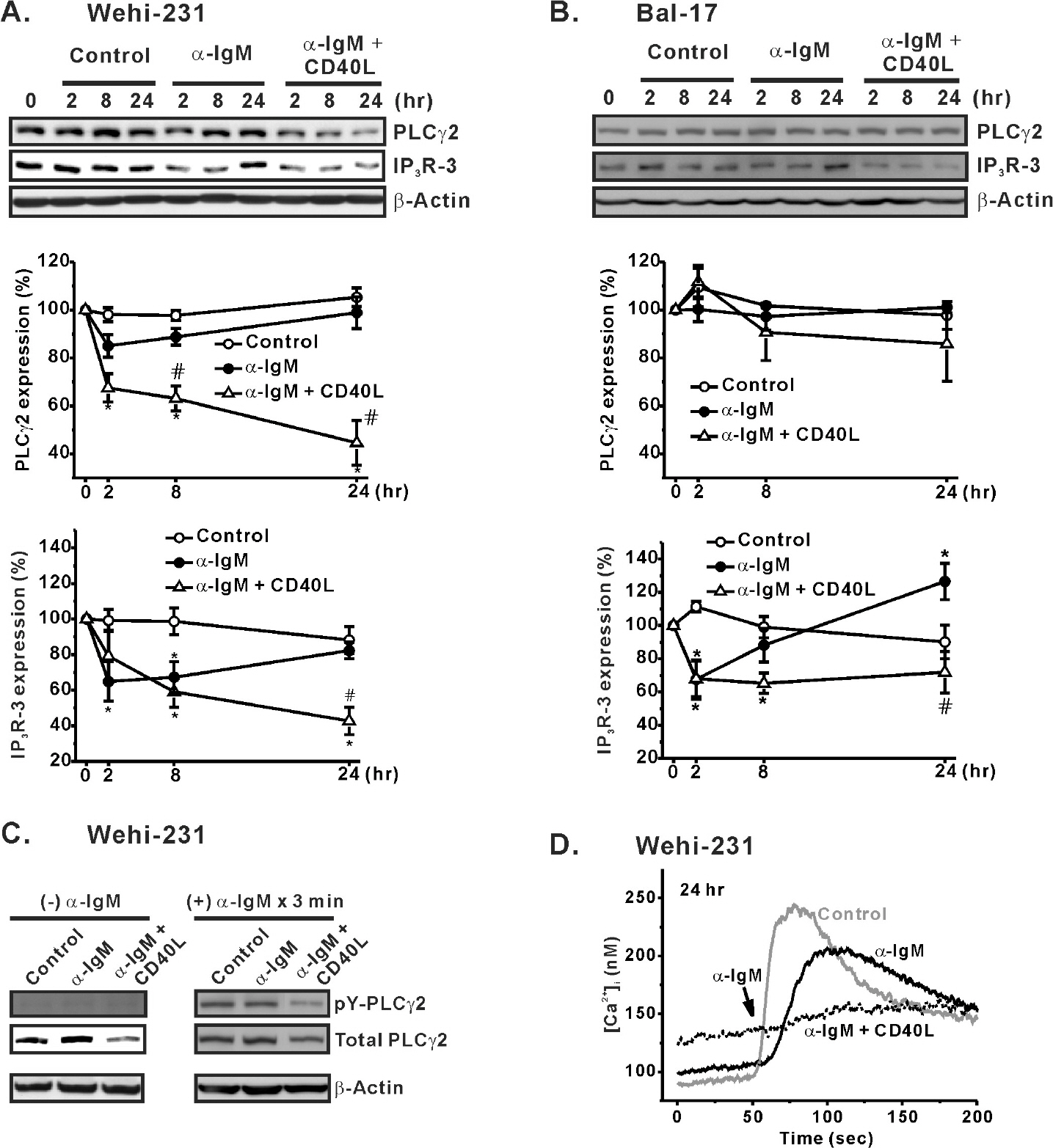
Fig. 5. Effects of CD40 co-stimulation on expression of PLCγ2 and IP3R-3. WEHI-231 (A) and Bal-17 (B) cells were treated long-term with α-IgM in the absence or presence of CD40 co-stimulation. Extracted total protein content was subjected to Western blotting to measure PLCγ2 and IP3R-3 expression. β-Actin was used as a protein loading control. The quantified intensities of PLCγ2 (PLC32/β-actin) and IP3R-3 (IP3R-3/β-actin) were normalized (% expression) to those of un-treated cells and then plotted against the indicated times of stimulation. Mean±S.E.M. of three independent experiments are shown in A and B. ∗p<0.05 compared to untreated cells. #p<0.05 compared to α-IgM-treated cells. (C) WEHI-231 cells were treated for 24 hr with α-IgM or α-IgM + CD40L. The harvested cells were re-suspended in NT solution and stimulated with α-IgM (5 μg/ml) for 3 min at 37°C. The α-IgM-stimulated cells were immunoprecipitated to measure the amount of tyrosine-phosphorylated PLCγ2 (pY-PLCγ2). Densities of pY-PLCγ2 were compared against those of total PLCγ2 and β-actin. (D) The amounts of pY-PLCγ2 in response to 3 min of BCR ligation in C were clearly determined based on the amplitudes of α-IgM-triggered [Ca2+]i increases in WEHI-231 cells.
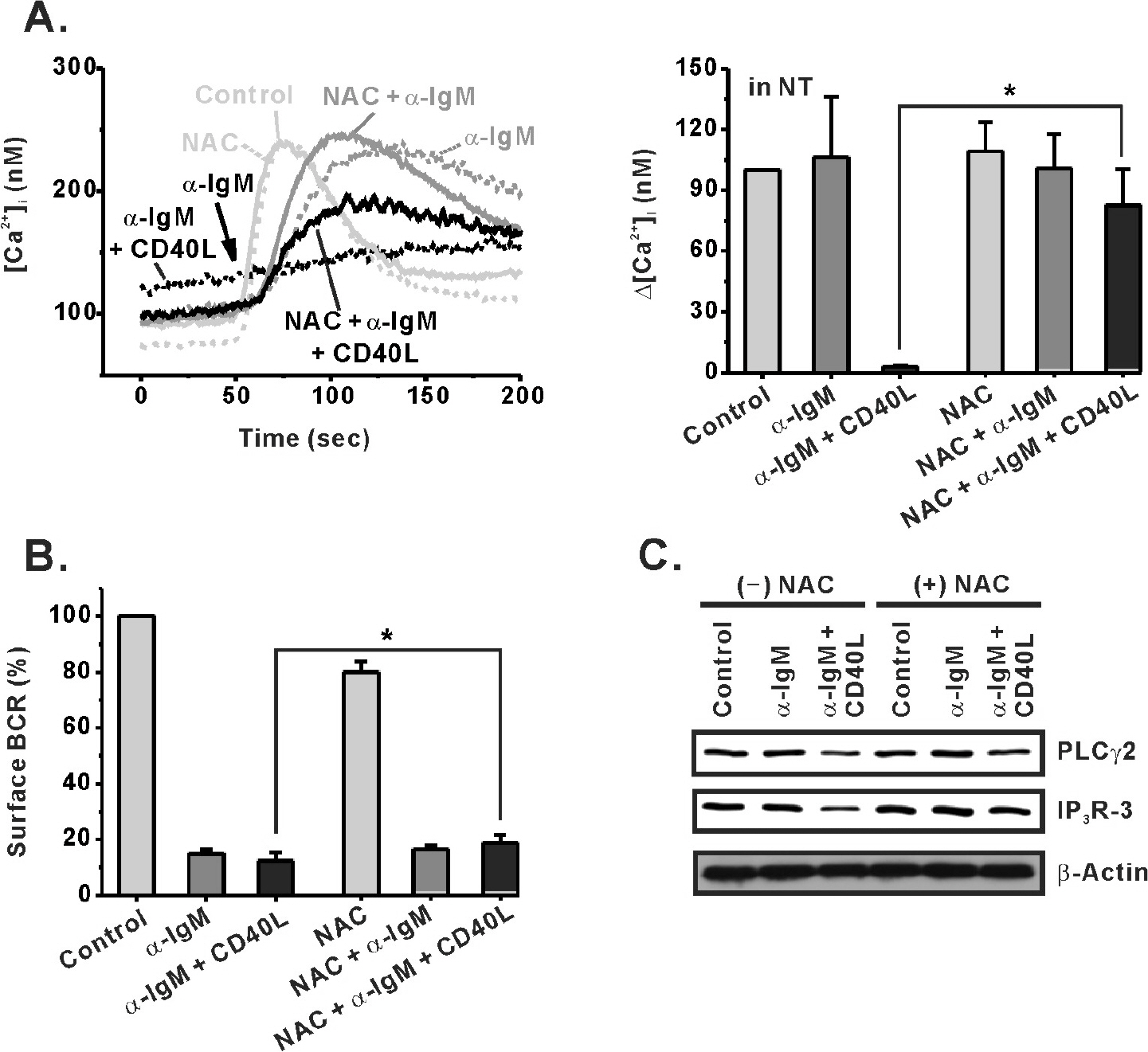
Fig. 6. An antioxidant, NAC, inhibits the effect of CD40 co-stimulation in WEHI-231 cells. An antioxidant, NAC (10 mM), was treated 30 min before stimulation of WEHI-231 cells with α-IgM or α-IgM + CD40L. After 24 hr of stimulation in the absence or presence of NAC, the cells were harvested and subjected to measurement of BCR-induced [Ca2+]i increase in NT solution (A), analysis of surface BCR level (B), and Western blotting for PLCγ2 and IP3R-3 expression (C). Mean±S.E.M. ∗p<0.05.
Reference
-
References
1. Niiro H, Clark EA. Regulation of B-cell fate by antigen-receptor signals. Nat Rev Immunol. 2002; 2:945–956.
Article2. Scharenberg AM, Humphries LA, Rawlings DJ. Calcium signalling and cell-fate choice in B cells. Nat Rev Immunol. 2007; 7:778–789.
Article3. Koncz G, Bodor C, Kövesdi D, Gáti R, Sármay G. BCR mediated signal transduction in immature and mature B cells. Immunol Lett. 2002; 82:41–49.
Article4. Harwood NE, Batista FD. New insights into the early molecular events underlying B cell activation. Immunity. 2008; 28:609–619.
Article5. Donjerković D, Zhang L, Scott DW. Regulation of p27Kip1 accumulation in murine B-lymphoma cells: role of c-Myc and calcium. Cell Growth Differ. 1999; 10:695–704.6. Chakraborti T, Das S, Mondal M, Roychoudhury S, Chakraborti S. Oxidant, mitochondria and calcium: an overview. Cell Signal. 1999; 11:77–85.7. Katz E, Deehan MR, Seatter S, Lord C, Sturrock RD, Harnett MM. B cell receptor-stimulated mitochondrial phospholipase A2 activation and resultant disruption of mitochondrial membrane potential correlate with the induction of apoptosis in WEHI-231 B cells. J Immunol. 2001; 166:137–147.8. van Kooten C, Banchereau J. CD40-CD40 ligand. J Leukoc Biol. 2000; 67:2–17.
Article9. Katz E, Lord C, Ford CA, Gauld SB, Carter NA, Harnett MM. Bcl-(xL) antagonism of BCR-coupled mitochondrial phospholipase A(2) signaling correlates with protection from apoptosis in WEHI-231 B cells. Blood. 2004; 103:168–176.
Article10. Lee HH, Dadgostar H, Cheng Q, Shu J, Cheng G. NF-kappaB-mediated up-regulation of Bcl-x and Bfl-1/A1 is required for CD40 survival signaling in B lymphocytes. Proc Natl Acad Sci USA. 1999; 96:9136–9141.11. Mineva ND, Rothstein TL, Meyers JA, Lerner A, Sonenshein GE. CD40 ligand-mediated activation of the de novo RelB NF-kappaB synthesis pathway in transformed B cells promotes rescue from apoptosis. J Biol Chem. 2007; 282:17475–17485.12. Watanabe K, Ichinose S, Hayashizaki K, Tsubata T. Induction of autophagy by B cell antigen receptor stimulation and its inhibition by costimulation. Biochem Biophys Res Commun. 2008; 374:274–281.
Article13. Yan BC, Adachi T, Tsubata T. ER stress is involved in B cell antigen receptor ligation-induced apoptosis. Biochem Biophys Res Commun. 2008; 365:143–148.
Article14. Lee JR. Reactive oxygen species play roles on B cell surface receptor CD40-mediated proximal and distal signaling events: effects of an antioxidant, N-acetyl-L-cysteine treatment. Mol Cell Biochem. 2003; 252:1–7.15. Tsien RY. New tetracarboxylate chelators for fluorescence measurement and photochemical manipulation of cytosolic free calcium concentrations. Soc Gen Physiol Ser. 1986; 40:327–345.16. Malgaroli A, Milani D, Meldolesi J, Pozzan T. Fura-2 measurement of cytosolic free Ca2+ in monolayers and suspensions of various types of animal cells. J Cell Biol. 1987; 105:2145–2155.17. Nunez R. DNA measurement and cell cycle analysis by flow cytometry. Curr Issues Mol Biol. 2001; 3:67–70.18. Riccardi C, Nicoletti I. Analysis of apoptosis by propidium iodide staining and flow cytometry. Nat Protoc. 2006; 1:1458–1461.
Article19. Khan AA, Soloski MJ, Sharp AH, Schilling G, Sabatini DM, Li SH, Ross CA, Snyder SH. Lymphocyte apoptosis: mediation by increased type 3 inositol 1,4,5-trisphosphate receptor. Science. 1996; 273:503–507.
Article20. Benschop RJ, Brandl E, Chan AC, Cambier JC. Unique signaling properties of B cell antigen receptor in mature and immature B cells: implications for tolerance and activation. J Immunol. 2001; 167:4172–4179.
Article21. Lee YJ, Kim NY, Suh YA, Lee C. Involvement of ROS in curcumin-induced autophagic cell death. Korean J Physiol Pharmacol. 2011; 15:1–7.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Differential Signaling via Tumor Necrosis Factor-Associated Factors (TRAFs) by CD27 and CD40 in Mouse B Cells
- Role for CD40 and CD40L Expression in Generating CD8 T Cell Response to Minor Histcompatibility Antigen, H60
- Ca2+-dependent Long-term Inactivation of Cardiac Na+/Ca2+ Exchanger
- Expression of Ca2+-dependent Synaptotagmin Isoforms in Mouse and Rat Parotid Acinar Cells
- Role of Regulators of G-Protein Signaling 4 in Ca2+ Signaling in Mouse Pancreatic Acinar Cells
