Swiprosin-1 Expression Is Up-Regulated through Protein Kinase C-theta and NF-kappaB Pathway in T Cells
- Affiliations
-
- 1School of Life Sciences, Immune Synapse Research Center and Cell Dynamics Research Center, Gwangju Institute of Science and Technology, Gwangju 500-712, Korea. cdjun@gist.ac.kr
- KMID: 1431926
- DOI: http://doi.org/10.4110/in.2013.13.2.55
Abstract
- Swiprosin-1 exhibits the highest expression in CD8+ T cells and immature B cells and has been proposed to play a role in lymphocyte biology through actin remodeling. However, regulation of swiprosin-1 gene expression is poorly understood. Here we report that swiprosin-1 is up-regulated in T cells by PKC pathway. Targeted inhibition of the specific protein kinase C (PKC) isotypes by siRNA revealed that PKC-theta is involved in the expression of swiprosin-1 in the human T cells. In contrast, down-regulation of swiprosin-1 by A23187 or ionomycin suggests that calcium-signaling plays a negative role. Interestingly, swiprosin-1 expression is only reduced by treatment with NF-kappaB inhibitors but not by NF-AT inhibitor, suggesting that the NF-kappaB pathway is critical for regulation of swiprosin-1 expression. Collectively, these results suggest that swiprosin-1 is a PKC-theta-inducible gene and that it may modulate the late phase of T cell activation after antigen challenge.
Keyword
MeSH Terms
Figure
-
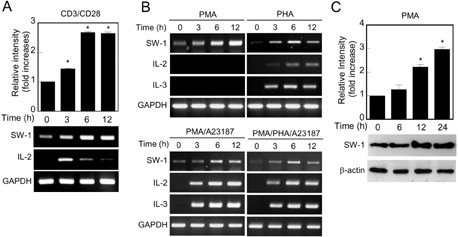
Figure 1 Expression analysis of swiprosin-1 mRNA by RT-PCR. (A) Expression of swiprosin-1 mRNA in Jurkat T cells. Jurkat T cells were stimulated with anti-CD3/CD28, and then the cells were harvested at the indicated time points (0~12 h). The mRNA levels of swiprosin-1 were determined by realtime quantitative PCR (top) and RT-PCR (bottom). GAPDH was used as an internal control. *p<0.01 versus 0 h. (B) Induction of swiprosin-1 in T cells by various stimuli. Jurkat T cells were treated with PMA (200 nM), PHA (1µg/ml), PMA/A23187 (1µM), or PMA/PHA/A23187. At the indicated time points, the expressions of swiprosin-1, IL-2, and IL-3 were determined by RT-PCR. GAPDH was used as an internal control. (C) Western blot analysis of swiprosin-1 expression in T cells. Jurkat T cells were activated by PMA, and then the cells were harvested at the indicated time points (0~24 h). The protein levels were determined by Western blot. All data are from at least three separate experiments. *p<0.01 versus 0 h.
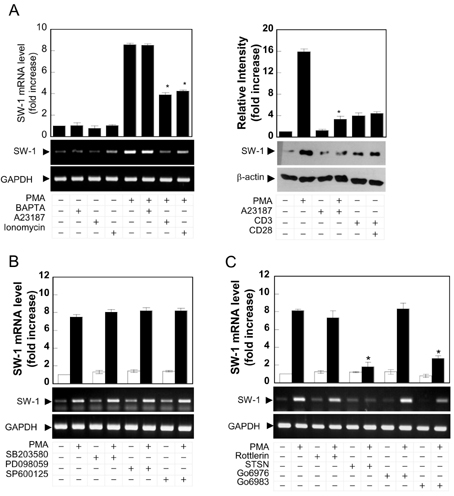
Figure 2 Swiprosin-1 is up-regulated by PMA but down-regulated by calcium ionophore. (A) Swiprosin-1 is regulated by PKC and calcium modulators. Jurkat T cells were pretreated for 30 min with BAPTAAM (50µM), A23187, or ionomycin (1µM), and then the cells were further incubated with medium alone or PMA, anti-CD3 (10µg/ml), and/or anti-CD28 (2µg/ml). After 6 h of incubation, the mRNA levels were determined as described in Fig. 1A. The protein levels were determined at the 18 h as described in Fig. 1C. Data represent three separate experiments. The data shown in the bar graphs represent mean±SD values of triplicate experiments. *p<0.01 versus PMA alone. (B) Jurkat T cells were pretreated for 30 min with MAP kinase inhibitors including SB203580 (10µM), PD098059 (10µM), and SP600125 (10µM), and then the cells were further stimulated for 6 h with PMA. The mRNA levels were determined as described in Fig. 1A. (C) Jurkat T cells were pretreated for 30 min with PKC inhibitors including Rottlerin (10µM), staurosporine (STSN, 500 nM), Gö6976 (100 nM), and Gö6983 (100 nM), and then the cells were further stimulated for 6 h with PMA. The mRNA levels were determined as described in Fig. 1A. Data represent three separate experiments. *p<0.01 versus PMA alone.

Figure 3 Swiprosin-1 expression is dependent on PKC-θ pathways in T cells. (A) Detection of PKC isoforms by Western blot in Jurkat T cells. (B) Knock-down of PKC isoforms by specific siRNA. Jurkat T cells were transfected with siRNA targeting specific PKC isoforms. At the indicated time points, the expression of PKC isoforms was analyzed by Western blot. Blots were also probed with antibody to β-actin to confirm an equal loading. (C) Knock-down of PKC-θ inhibits phorbol ester-induced swiprosin-1 expression in Jurkat T cells. After 72 h of siRNA transfection against indicated PKC isoforms, cells were treated with or without PMA (200 nM) for 6 h and the expression of swiprosin-1 was analyzed by RT-PCR (top) and quantitative RT-PCR (bottom). Amplification of GAPDH was used as an internal control. Data represent three separate experiments. The data shown in the bar graphs represent mean±SD values of triplicate experiments. *p<0.01 versus scrambled siRNA. SC, scrambled.

Figure 4 Swiprosin-1 expression is regulated by the PKC-θ pathway in Jurkat T and human primary PBLs. (A and B) Jurkat T cells (A) and primary PBLs (B) were stimulated with anti-CD3/CD28, and then the cells were harvested and the mRNA levels were determined as described by Fig. 1A. The expression of PKC isoforms in primary T cells was analyzed by Western blot. Data represent three separate experiments. The data shown in the bar graphs represent mean±SD values of triplicate experiments. *p<0.01 versus scrambled siRNA. SC, scrambled.

Figure 5 Swiprosin-1 expression is regulated by the NF-κB but not by the NF-AT. (A) Jurkat T cells were transiently transfected with NF-κB or NF-AT luciferase reporter constructs. After 24 h, the cells were treated with PMA, A23187, or PMA/A23187 and incubated for 12 h. NF-κB- or NF-AT-dependent transcriptional activities were determined by luciferase activity assay. Data are representative of three separate experiments. The data shown in bar graphs represent mean±SD values of triplicate experiments. (B) Jurkat T cells were pretreated for 30 min with NF-κB inhibitors including MG132 (1µM), and CAPE (10µM) or NF-AT inhibitor CsA (1µg/ml), and then the cells were further stimulated for 6 h with PMA (left) or PMA/A23187 (right). The mRNA levels were determined as described in Fig. 1A. Data represent three separate experiments.
Reference
-
1. Vuadens F, Rufer N, Kress A, Corthésy P, Schneider P, Tissot JD. Identification of swiprosin 1 in human lymphocytes. Proteomics. 2004. 4:2216–2220.
Article2. Avramidou A, Kroczek C, Lang C, Schuh W, Jäck HM, Mielenz D. The novel adaptor protein Swiprosin-1 enhances BCR signals and contributes to BCR-induced apoptosis. Cell Death Differ. 2007. 14:1936–1947.
Article3. Ramesh TP, Kim YD, Kwon MS, Jun CD, Kim SW. Swiprosin-1 regulates cytokine expression of human mast cell line HMC-1 through actin remodeling. Immune Netw. 2009. 9:274–284.
Article4. Blagoev B, Ong SE, Kratchmarova I, Mann M. Temporal analysis of phosphotyrosine-dependent signaling networks by quantitative proteomics. Nat Biotechnol. 2004. 22:1139–1145.
Article5. Mielenz D, Vettermann C, Hampel M, Lang C, Avramidou A, Karas M, Jäck HM. Lipid rafts associate with intracellular B cell receptors and exhibit a B cell stage-specific protein composition. J Immunol. 2005. 174:3508–3517.
Article6. Piragyte I, Jun CD. Actin engine in immunological synapse. Immune Netw. 2012. 12:71–83.
Article7. Meng X, Wilkins JA. Compositional characterization of the cytoskeleton of NK-like cells. J Proteome Res. 2005. 4:2081–2087.
Article8. Thylur RP, Kim YD, Kwon MS, Oh HM, Kwon HK, Kim SH, Im SH, Chun JS, Park ZY, Jun CD. Swiprosin-1 is expressed in mast cells and up-regulated through the protein kinase C beta I/eta pathway. J Cell Biochem. 2009. 108:705–715.
Article9. Hayashi K, Altman A. Protein kinase C theta (PKCtheta): a key player in T cell life and death. Pharmacol Res. 2007. 55:537–544.10. Freeley M, Long A. Regulating the regulator: Phosphorylation of PKC θ in T Cells. Front Immunol. 2012. 3:227.
Article11. Kwon MJ, Wang R, Ma J, Sun Z. PKC-θ is a drug target for prevention of T cell-mediated autoimmunity and allograft rejection. Endocr Metab Immune Disord Drug Targets. 2010. 10:367–372.
Article12. Manicassamy S, Yin D, Zhang Z, Molinero LL, Alegre ML, Sun Z. A critical role for protein kinase C-theta-mediated T cell survival in cardiac allograft rejection. J Immunol. 2008. 181:513–520.
Article13. Murphy KM, Reiner SL. The lineage decisions of helper T cells. Nat Rev Immunol. 2002. 2:933–944.
Article14. Shahabi NA, McAllen K, Sharp BM. Stromal cell-derived factor 1-alpha (SDF)-induced human T cell chemotaxis becomes phosphoinositide 3-kinase (PI3K)-independent: role of PKC-theta. J Leukoc Biol. 2008. 83:663–671.
Article15. Tan SL, Parker PJ. Emerging and diverse roles of protein kinase C in immune cell signalling. Biochem J. 2003. 376:545–552.
Article16. Michalczyk I, Sikorski AF, Kotula L, Junghans RP, Dubielecka PM. The emerging role of protein kinase Cθ in cytoskeletal signaling. J Leukoc Biol. 2013. 93:319–327.
Article17. Manicassamy S, Gupta S, Huang Z, Sun Z. Protein kinase C-theta-mediated signals enhance CD4XMLLink_XYZ T cell survival by up-regulating Bcl-xL. J Immunol. 2006. 176:6709–6716.
Article18. Isakov N, Altman A. Protein kinase C(theta) in T cell activation. Annu Rev Immunol. 2002. 20:761–794.19. Altman A, Villalba M. Protein kinase C-theta (PKC theta): a key enzyme in T cell life and death. J Biochem. 2002. 132:841–846.20. Boschelli DH. Small molecule inhibitors of PKCTheta as potential antiinflammatory therapeutics. Curr Top Med Chem. 2009. 9:640–654.
Article21. Lanzavecchia A. Understanding the mechanisms of sustained signaling and T cell activation. J Exp Med. 1997. 185:1717–1719.
Article22. Gardner P. Calcium and T lymphocyte activation. Cell. 1989. 59:15–20.
Article23. Rao A, Luo C, Hogan PG. Transcription factors of the NFAT family: regulation and function. Annu Rev Immunol. 1997. 15:707–747.24. Chan AC, Desai DM, Weiss A. The role of protein tyrosine kinases and protein tyrosine phosphatases in T cell antigen receptor signal transduction. Annu Rev Immunol. 1994. 12:555–592.
Article25. Bootman MD, Berridge MJ. The elemental principles of calcium signaling. Cell. 1995. 83:675–678.
Article26. Vega IE, Traverso EE, Ferrer-Acosta Y, Matos E, Colon M, Gonzalez J, Dickson D, Hutton M, Lewis J, Yen SH. A novel calcium-binding protein is associated with tau proteins in tauopathy. J Neurochem. 2008. 106:96–106.
Article27. Dütting S, Brachs S, Mielenz D. Fraternal twins: Swiprosin-1/EFhd2 and Swiprosin-2/EFhd1, two homologous EF-hand containing calcium binding adaptor proteins with distinct functions. Cell Commun Signal. 2011. 9:2.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Swiprosin-1 Regulates Cytokine Expression of Human Mast Cell Line HMC-1 through Actin Remodeling
- PMA-induced up-regulation of MMP-9 is regulated by a PKCalpha-NF-kappaB cascade in human lung epithelial cells
- Role of protein kinases on NF- kappaB activation and cell death in bovine cerebral endothelial cells
- ASK1 is Involved in EBV LMP1-induced NF-kappaB Activation
- NF-kappaB Activation in T Helper 17 Cell Differentiation
