Resveratrol Inhibits GABAC rho Receptor-Mediated Ion Currents Expressed in Xenopus Oocytes
- Affiliations
-
- 1Department of Physiology, College of Veterinary Medicine and Bio-Molecular Informatics Center, Konkuk University, Seoul 143-701, Korea. synah@konkuk.ac.kr
- 2Department of Physical Therapy, Sehan University, Yeongam 526-702, Korea.
- KMID: 1429390
- DOI: http://doi.org/10.4196/kjpp.2013.17.2.175
Abstract
- Resveratrol is a phytoalexin found in grapes, red wine, and berries. Resveratrol has been known to have many beneficial health effects, such as anti-cancer, neuroprotective, anti-inflammatory, and life-prolonging effects. However, relatively little is known about the effects of resveratrol on the regulation of ligand-gated ion channels. We have previously reported that resveratrol regulates subsets of homomeric ligand-gated ion channels such as those of 5-HT3A receptors. The gamma-aminobutyric acidC (GABAC) receptor is mainly expressed in retinal bipolar cells and plays an important role in visual processing. In the present study, we examined the effects of resveratrol on the channel activity of homomeric GABAC receptor expressed in Xenopus oocytes injected with cRNA encoding human GABAC rho subunits. Our data show that the application of GABA elicits an inward peak current (IGABA) in oocytes that express the GABAC receptor. Resveratrol treatment had no effect on oocytes injected with H2O or with GABAC receptor cRNA. Co-treatment with resveratrol and GABA inhibited IGABA in oocytes with GABAC receptors. The inhibition of IGABA by resveratrol was in a reversible and concentration-dependent manner. The IC50 of resveratrol was 28.9+/-2.8 microM in oocytes expressing GABAC receptor. The inhibition of IGABA by resveratrol was in voltage-independent and non-competitive manner. These results indicate that resveratrol might regulate GABAC receptor expression and that this regulation might be one of the pharmacological actions of resveratrol on the nervous system.
MeSH Terms
-
Fruit
gamma-Aminobutyric Acid
Humans
Inhibitory Concentration 50
Ligand-Gated Ion Channels
Nervous System
Oocytes
Receptors, GABA
Retinal Bipolar Cells
RNA, Complementary
Sesquiterpenes
Stilbenes
Vitis
Wine
Xenopus
Ligand-Gated Ion Channels
RNA, Complementary
Receptors, GABA
Sesquiterpenes
Stilbenes
gamma-Aminobutyric Acid
Figure
-
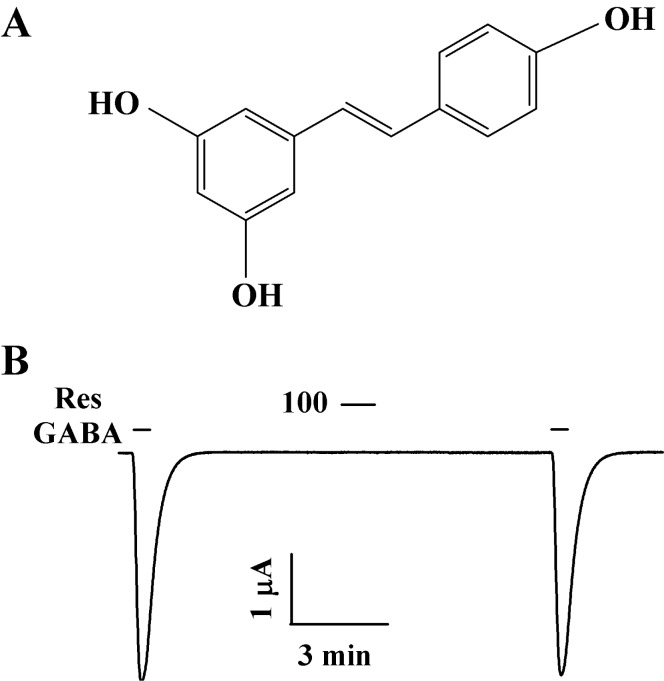
Fig. 1 Chemical structure of resveratrol and effect of resveratrol (Res) on GABAC receptors expressing oocytes. (A). Application of resveratrol (100 µM) for 1 min had no effect on IGABA expression in oocytes expressing GABAC receptors (B).
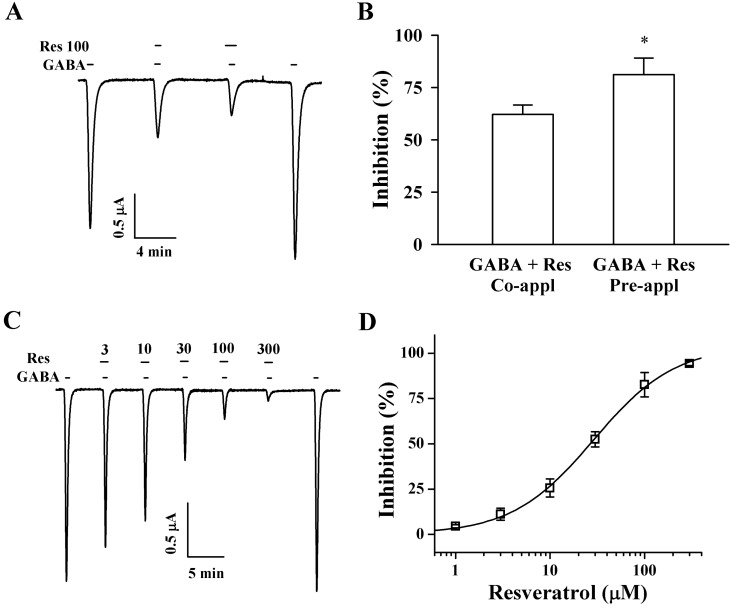
Fig. 2 Effect of Res on IGABA expression in oocytes that express GABAC receptors. (A) GABA (2 µM) was first applied and then GABA was co- or pre-applied with Res (100 µM). Thus, co- and pre-application of Res with GABA inhibited IGABA. The resting membrane potential of oocytes was approximately -35 mV, and oocytes were voltage clamped at a holding potential of -80 mV prior to drug application. Traces are representative of 6 separate oocytes from 3 frogs. (B) Pre-application of Res inhibited IGABA more potently than that inhibited by co-treatment. (C) IGABA in GABAC receptors expressing oocytes was elicited at -80 mV holding potential, with indicated time in the presence of 2 µM GABA and with the indicated pre-treatment concentration of Res that was applied before GABA application. (D) IGABA % inhibition induced by Res treatment was calculated using the average of peak that the inward current elicited by GABA treatment before Res application and of the peak inward current elicited by GABA treatment after pre-treatment of Res before GABA. The continuous line shows the curve fitted according to the equation. Each point represents the mean±S.E.M. (n=9~12 from 3 frogs).
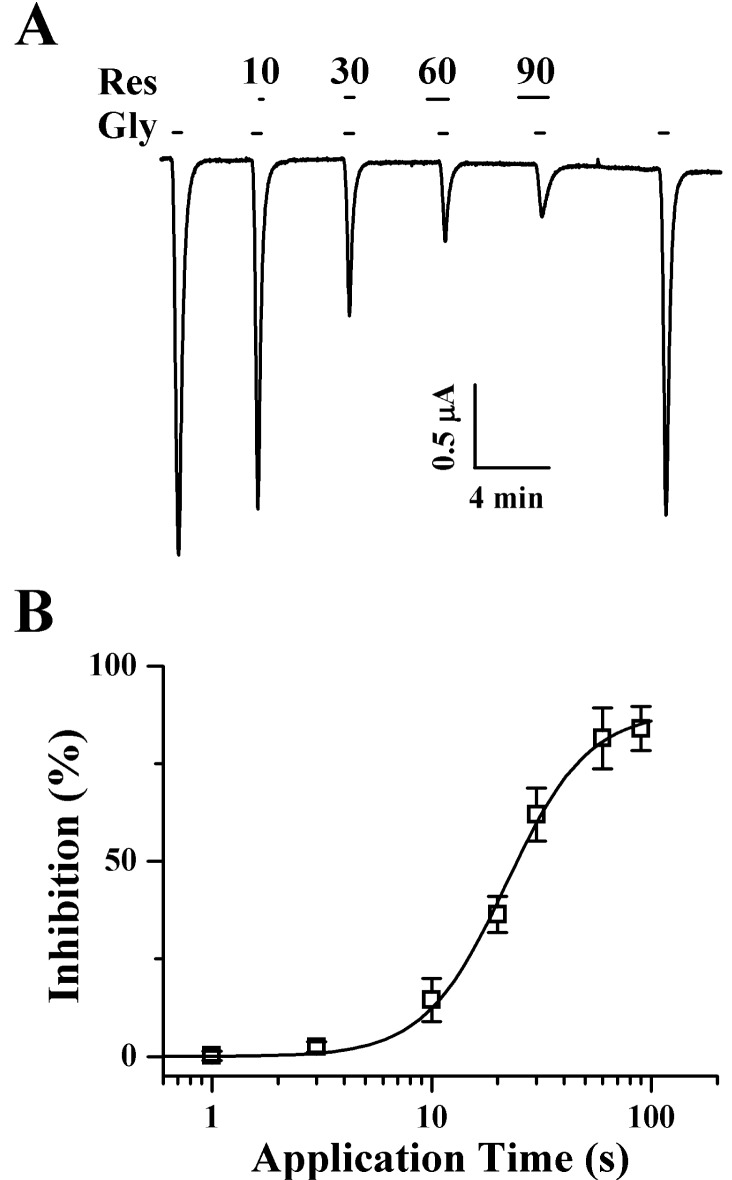
Fig. 3 Time-dependent effects of pre-application of Res on IGABA in oocytes that express GABAC receptors. (A) Res (100 µM)-mediated inhibition on IGABA is pre-application-time dependent. Traces represent 6 separate oocytes from 3 frog batches. IGABA in GABAC receptor-expressing oocytes was elicited at a holding potential of -80 mV for the indicated Res pre-application time prior to drug application. (B) Res-mediated inhibition of IGABA was almost saturated after 30 s of pre-application. The resting membrane potential of the oocytes was approximately -35 mV, and the oocytes were voltage-clamped at a holding potential of -80 mV. Each point represents the mean±S.E.M. (n=9~12/group).
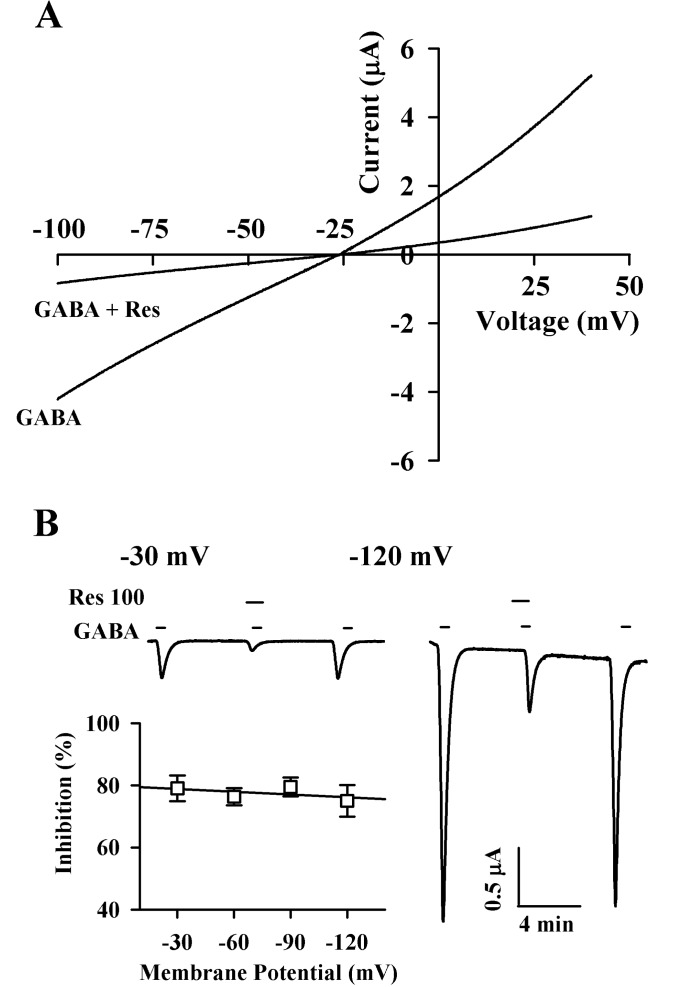
Fig. 4 Current-voltage relationship and voltage-independent inhibition by Res. (A) Current-voltage relationships of IGABA inhibition by Res in GABAC receptor-expressing oocytes. Representative current-voltage relationships were obtained using voltage ramps of -100 to +40 mV for 300 ms at a holding potential of -80 mV. Voltage steps were applied before and after application of 2 µM GABA in the absence or presence of 100 µM Res. (B) Voltage-independent inhibition of IGABA in the GABAC receptors by Res. Inset; the values were obtained from the receptors in the presence or absence of 100 µM Res at the indicated membrane holding potentials.
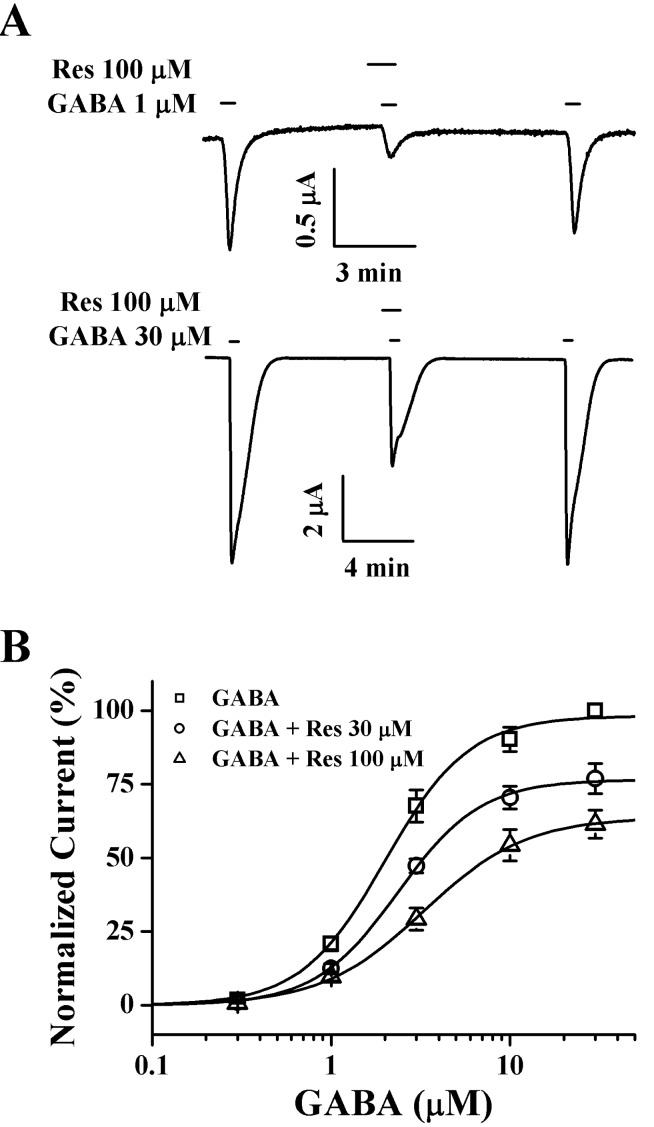
Fig. 5 Concentration-dependent effects of GABA on Res-mediated inhibition of IGABA. (A) The representative traces were obtained from the GABAC receptor-expressing oocytes. IGABA expression shown in the upper and lower panels were elicited at a holding potential of -80 mV by GABA at concentrations of 1 µM and 30 µM GABA respectively. (B) Concentration-response relationship of GABA with GABAC receptors treated with GABA (0.3~30 µM) alone or with GABA plus pre-application of 30 µM or 100 µM Res. The IGABA of oocytes expressing the GABAC receptors was measured using the indicated concentration of GABA in the absence (□) or presence of 30 µM (○) or 100 µM (▵) Res. Oocytes were exposed to GABA alone or to GABA with Res. Oocytes were voltage-clamped at a holding potential of -80 mV. Each point represents mean±S.E.M. (n=9~12/group).
Reference
-
1. Jensen ML, Schousboe A, Ahring PK. Charge selectivity of the Cys-loop family of ligand-gated ion channels. J Neurochem. 2005; 92:217–225. PMID: 15663470.
Article2. Bormann J. The 'ABC' of GABA receptors. Trends Pharmacol Sci. 2000; 21:16–19. PMID: 10637650.
Article3. Chebib M, Johnston GA. GABA-Activated ligand gated ion channels: medicinal chemistry and molecular biology. J Med Chem. 2000; 43:1427–1447. PMID: 10780899.
Article4. Milligan CJ, Buckley NJ, Garret M, Deuchars J, Deuchars SA. Evidence for inhibition mediated by coassembly of GABAA and GABAC receptor subunits in native central neurons. J Neurosci. 2004; 24:7241–7250. PMID: 15317850.5. Bloom FE, Iversen LL. Localizing 3H-GABA in nerve terminals of rat cerebral cortex by electron microscopic autoradiography. Nature. 1971; 229:628–630. PMID: 4925465.6. McCabe RT, Wamsley JK. Autoradiographic localization of subcomponents of the macromolecular GABA receptor complex. Life Sci. 1986; 39:1937–1945. PMID: 3023773.7. Wässle H, Koulen P, Brandstätter JH, Fletcher EL, Becker CM. Glycine and GABA receptors in the mammalian retina. Vision Res. 1998; 38:1411–1430. PMID: 9667008.
Article8. McCall MA, Lukasiewicz PD, Gregg RG, Peachey NS. Elimination of the rho1 subunit abolishes GABA(C) receptor expression and alters visual processing in the mouse retina. J Neurosci. 2002; 22:4163–4174. PMID: 12019334.9. Drew CA, Johnston GA, Weatherby RP. Bicuculline-insensitive GABA receptors: studies on the binding of (-)-baclofen to rat cerebellar membranes. Neurosci Lett. 1984; 52:317–321. PMID: 6097844.
Article10. Strata F, Cherubini E. Transient expression of a novel type of GABA response in rat CA3 hippocampal neurones during development. J Physiol. 1994; 480:493–503. PMID: 7869263.
Article11. Johnston GA, Chebib M, Hanrahan JR, Mewett KN. GABA(C) receptors as drug targets. Curr Drug Targets CNS Neurol Disord. 2003; 2:260–268. PMID: 12871036.
Article12. Langcake P, Pryce RJ. A new class of phytoalexins from grapevines. Experientia. 1977; 33:151–152. PMID: 844529.
Article13. Dudley J, Das S, Mukherjee S, Das DK. Resveratrol, a unique phytoalexin present in red wine, delivers either survival signal or death signal to the ischemic myocardium depending on dose. J Nutr Biochem. 2009; 20:443–452. PMID: 18789672.14. Pervaiz S. Resveratrol: from grapevines to mammalian biology. FASEB J. 2003; 17:1975–1985. PMID: 14597667.
Article15. Valenzano DR, Terzibasi E, Genade T, Cattaneo A, Domenici L, Cellerino A. Resveratrol prolongs lifespan and retards the onset of age-related markers in a short-lived vertebrate. Curr Biol. 2006; 16:296–300. PMID: 16461283.
Article16. Chen J, Zhou Y, Mueller-Steiner S, Chen LF, Kwon H, Yi S, Mucke L, Gan L. SIRT1 protects against microglia-dependent amyloid-beta toxicity through inhibiting NF-kappaB signaling. J Biol Chem. 2005; 280:40364–40374. PMID: 16183991.17. West T, Atzeva M, Holtzman DM. Pomegranate polyphenols and resveratrol protect the neonatal brain against hypoxic-ischemic injury. Dev Neurosci. 2007; 29:363–372. PMID: 17762204.
Article18. Raval AP, Dave KR, Pérez-Pinzón MA. Resveratrol mimics ischemic preconditioning in the brain. J Cereb Blood Flow Metab. 2006; 26:1141–1147. PMID: 16395277.
Article19. Lee BH, Hwang SH, Choi SH, Shin TJ, Kang J, Lee SM, Nah SY. Resveratrol enhances 5-hydroxytryptamine type 3A receptor-mediated ion currents: the role of arginine 222 residue in pre-transmembrane domain I. Biol Pharm Bull. 2011; 34:523–527. PMID: 21467640.
Article20. Chan MM. Antimicrobial effect of resveratrol on dermatophytes and bacterial pathogens of the skin. Biochem Pharmacol. 2002; 63:99–104. PMID: 11841782.
Article21. Lee BH, Shin TJ, Hwang SH, Choi SH, Kang J, Kim HJ, Park CW, Lee SH, Nah SY. Inhibitory effects of quercetin on muscle-type of nicotinic acetylcholine receptor-mediated ion currents expressed in xenopus oocytes. Korean J Physiol Pharmacol. 2011; 15:195–201. PMID: 21994477.
Article22. Sine SM, Taylor P. Local anesthetics and histrionicotoxin are allosteric inhibitors of the acetylcholine receptor. Studies of clonal muscle cells. J Biol Chem. 1982; 257:8106–8114. PMID: 7085658.
Article23. Heidmann T, Oswald RE, Changeux JP. Multiple sites of action for noncompetitive blockers on acetylcholine receptor rich membrane fragments from torpedo marmorata. Biochemistry. 1983; 22:3112–3127. PMID: 6882740.
Article24. Arias HR. Luminal and non-luminal non-competitive inhibitor binding sites on the nicotinic acetylcholine receptor. Mol Membr Biol. 1996; 13:1–17. PMID: 9147657.
Article25. Saleh MC, Connell BJ, Saleh TM. Resveratrol preconditioning induces cellular stress proteins and is mediated via NMDA and estrogen receptors. Neuroscience. 2010; 166:445–454. PMID: 20040366.
Article26. Wu Z, Xu Q, Zhang L, Kong D, Ma R, Wang L. Protective effect of resveratrol againstkainate-induced temporal lobe epilepsy in rats. Neurochem Res. 2009; 34:1393–1400. PMID: 19219549.27. Shinohara Y, Toyohira Y, Ueno S, Liu M, Tsutsui M, Yanagihara N. Effects of resveratrol, a grape polyphenol, on catecholamine secretion and synthesis in cultured bovine adrenal medullary cells. Biochem Pharmacol. 2007; 74:1608–1618. PMID: 17888406.
Article28. Boue-Grabot E, Taupignon A, Tramu G, Garret M. Molecular and electrophysiological evidence for a GABAc receptor in thyrotropin-secreting cells. Endocrinology. 2000; 141:1627–1632. PMID: 10803570.29. Jansen A, Hoepfner M, Herzig KH, Riecken EO, Scherübl H. GABA(C) receptors in neuroendocrine gut cells: a new GABA-binding site in the gut. Pflugers Arch. 2000; 441:294–300. PMID: 11211116.
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Inhibitory Effects of Ginsenoside Metabolites, Compound K and Protopanaxatriol, on GABAC Receptor-Mediated Ion Currents
- Quercetin Inhibits alpha3beta4 Nicotinic Acetylcholine Receptor-Mediated Ion Currents Expressed in Xenopus Oocytes
- Block of ATP-sensitive K+ channels expressed in Xenopus oocytes by dimethyl sulfoxide
- Inhibitory Effects of Quercetin on Muscle-type of Nicotinic Acetylcholine Receptor-Mediated Ion Currents Expressed in Xenopus Oocytes
- Cloning of Xenopus laevis TRPV2 by Gene Prediction
