Effects of Microbiota on the Treatment of Obesity with the Natural Product Celastrol in Rats
- Affiliations
-
- 1State Key Laboratory of Reproductive Medicine, Center for Global Health, School of Public Health, Nanjing Medical University, Nanjing, China.
- 2Key Laboratory of Modern Toxicology of Ministry of Education, School of Public Health, Nanjing Medical University, Nanjing, China.
- KMID: 2513036
- DOI: http://doi.org/10.4093/dmj.2019.0124
Abstract
Background Obesity has become one of the most serious issues threatening the health of humankind, and we conducted this study to examine whether and how celastrol protects against obesity.
Methods We fed male Sprague-Dawley rats a high-fat diet and administered celastrol to obese rats for 3 weeks. By recording body weight (BW) and other measures, we identified the effective dose of celastrol for obesity treatment. Feces were collected to perform 16S rRNA sequencing, and hypothalami were extracted for transcriptome sequencing. We then treated leptin knockout rats with celastrol and explored the changes in energy metabolism. Male Institute of Cancer Research (ICR) mice were used to test the acute toxicity of celastrol.
Results We observed that celastrol reduced BW and promoted energy expenditure at a dose of 500 µg/kg BW but that food intake was not changed after administration. The diversity of the gut microbiota was improved, with an increased ratio of
Bacteroidetes toFirmicutes , and the gut microbiota played an important role in the anti-obesity effects of celastrol. Hypothalamic transcriptome analysis showed a significant enrichment of the leptin signaling pathway, and we found that celastrol significantly enhanced energy expenditure, which was mediated by the leptin signaling pathway. Acute lethal toxicity of celastrol was not observed at doses ranging from 0 to 62.5 mg/kg BW.Conclusion Our study revealed that celastrol decreased the BW of obese rats by enhancing energy expenditure but not by suppressing food intake and that this effect was mediated by the improvement of the gut microbiota and the activation of the hypothalamic leptin signaling pathway.
Figure
-
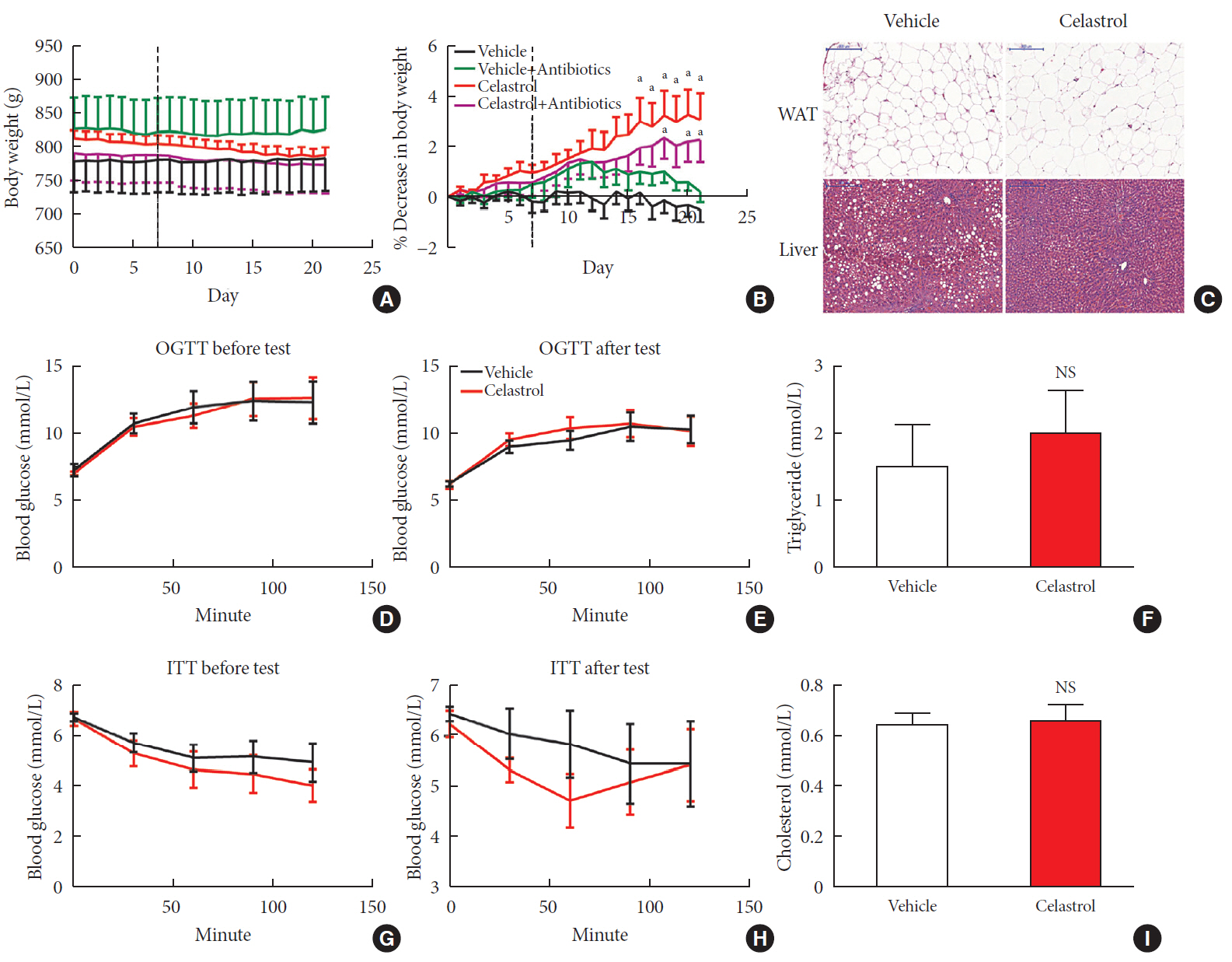
Fig. 1 Celastrol protects rats against diet-induced obesity. Diet-induced obese (DIO) Sprague-Dawley rats were orally administered vehicle or celastrol (500 µg/kg) every day for 3 weeks. (A) The body weight and (B) percent decrease (%) in the body weight of the DIO rats during the treatment period (n=8 for vehicle group; n=7 for celastrol group). (C) Representative H&E staining of abdominal white adipose tissue (WAT) and the liver. The results of the oral glucose tolerance test (OGTT) of the rats before (D) and after (E) the treatment period. (F) The serum triglyceride levels of the rats after 3 weeks of treatment. The results of the insulin tolerance test (ITT) of the rats before (G) and after (H) the treatment period. (I) The serum cholesterol levels of the rats after 3 weeks of treatment. The error bars represent the standard error of means. The P values were determined by (B) Dunnett's post hoc test or (D–I) Student's t-test. NS, not significant. aP<0.05.

Fig. 2 Celastrol alters the microbiota composition in diet-induced obese (DIO) rats. The microbiota composition of the feces of vehicle-, celastrol-, and celastrol+antibiotic-treated DIO rats was analyzed by 16S rRNA sequencing (n=6 for each group). (A) The weighted version of the UniFrac-based principal coordinate analysis (PCoA), (B) α-diversity, and (C) relative abundance (%) of the microbiota at the phylum level in vehicle- and celastrol-treated rats after 3 weeks of treatment. (D, E) The consecutive relative abundance of the microbiota of (D) all phyla, (E) Bacteroidetes (left panel), and Firmicutes (right panel) in vehicle- and celastrol-treated rats after 1, 2, and 3 weeks of treatment. (F) To understand the role of the microbiota in the anti-obesity effect of celastrol, antibiotics were added to the drinking water beginning the second week of celastrol treatment, as indicated by antibiotic intervention in the schematic diagram (n=7 for the celastrol group; n=8 for the antibiotic intervention group). (G) The α-diversity and (H) weighted version of the UniFrac-based PCoA of the microbiota in celastrol- and celastrol+antibiotic-treated rats after 3 weeks of treatment. (B, G) The P values were determined by the rank sum test. Alterations in the relative abundances of Bacteroidetes and Firmicutes were fitted by linear regression.

Fig. 3 Celastrol activates hypothalamic leptin signaling in diet-induced obese (DIO) rats without affecting serum leptin levels. (A) Principle component (PC) analysis of RNA-seq data from the hypothalami of rats after 3 weeks of treatment (n=3 for each group). (B) The hierarchical clustering of differentially expressed genes (left panel) and enriched pathways, as identified by ingenuity pathway analysis (right panel). (C) Western blot of the protein levels of downstream effectors of leptin, namely, phosphorylated-signal transducer and activator of transcription 3 (p-STAT3)Tyr705 and total STAT3, and β-actin, in the hypothalami of vehicle-, antibiotic-, celastrol-, and celastrol+antibiotic-treated rats. (D) The ratio of the signal intensity of p-STAT3 to that of total STAT3. (E) The serum leptin levels, as determined by enzyme-linked immunosorbent assay (ELISA), after celastrol treatment for 1, 2, and 3 weeks. (F) The mRNA expression of critical genes in the leptin signaling pathway, as determined by quantitative real-time polymerase chain reaction (RT-qPCR). The findings indicate that the activation of the leptin signaling pathway might result from an improvement in leptin sensitivity. The error bars represent the standard error of means. (E, F) The P values were determined by Student's t-test. DARPP32 (PPP1R1B), protein phosphatase 1 regulatory inhibitor subunit 1B; cAMP, cyclic adenosine monophosphate; GNRH, gonadotropin releasing hormone; CXCR4, C-X-C motif chemokine receptor 4; GABA, gamma-aminobutyric acid; nNOS, nitric oxide synthase; PPAR, peroxisome proliferator activated receptor; RXR, retinoid X receptor; ERK, extracellular signal-regulated kinases; MAPK, mitogen-activated protein kinases; ErbB (EGFR), epidermal growth factor receptor; PXR (NR1I2), nuclear receptor subfamily 1 group I member 2; IGF-1, insulin like growth factor 1; PI3K, phosphatidylinositol-4,5-bisphosphate 3-kinase; VDR, vitamin D receptor; NS, not significant; Agrp, agouti-related peptide; Npy, neuropeptide Y; Pomc, pro-opiomelanocortin; Ptpn1, protein tyrosine phosphatase non-receptor type 1; Socs3, suppressor of cytokine signaling 3.
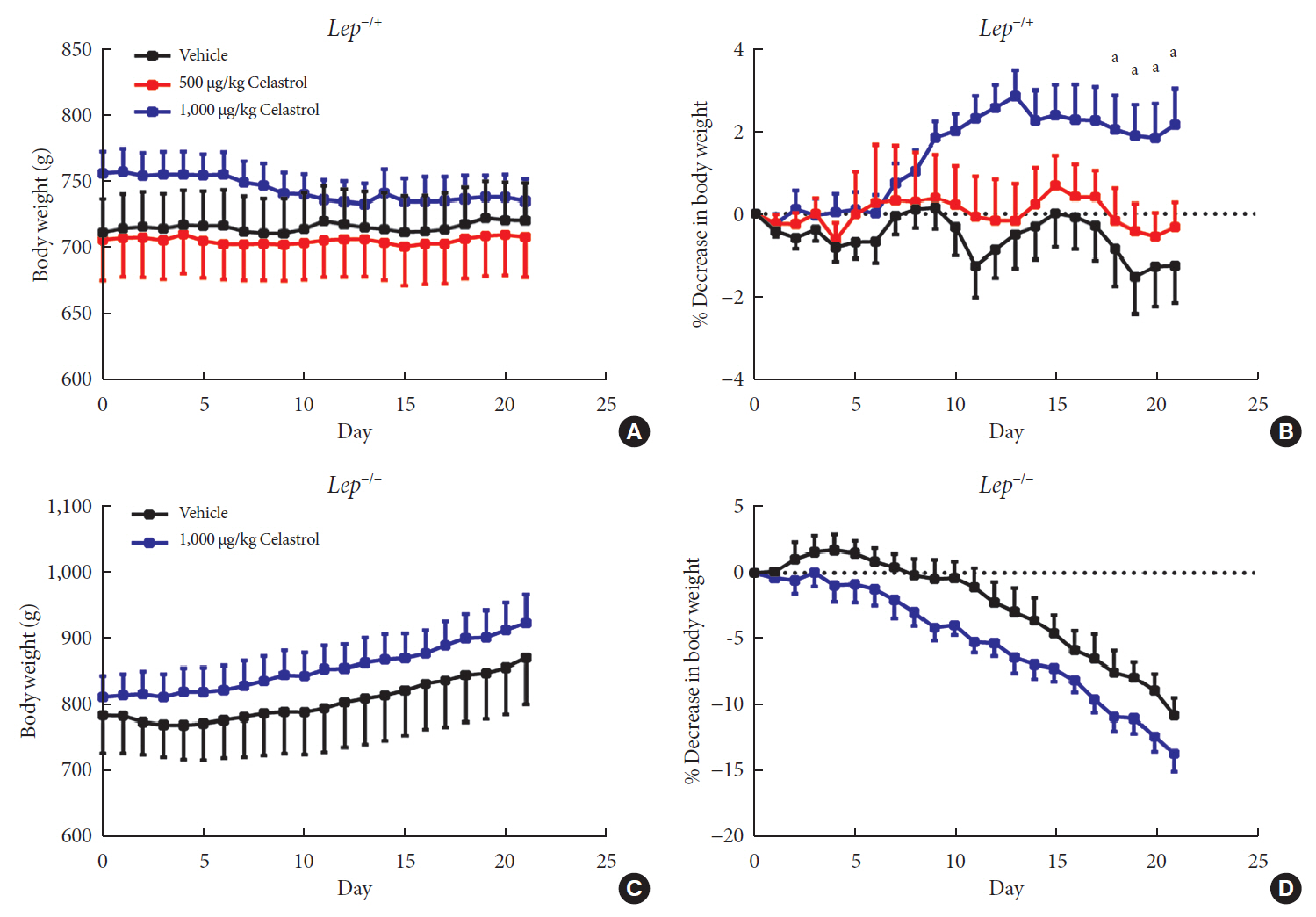
Fig. 4 A higher dose of celastrol protects Lep−/+ rats, but not Lep−/− rats, against high-fat-diet-induced obesity. Heterozygous (Lep−/+) and homozygous (Lep−/−) leptin knockout diet-induced obese rats were orally administered vehicle, 500 µg/kg celastrol or 1,000 µg/kg celastrol every day for 3 weeks. (A) The body weight and (B) the percent decrease (%) in body weight of the Lep−/+ rats during the treatment period (n=9 for the vehicle group; n=7 for the 500 µg/kg celastrol group; n=8 for the 1,000 µg/kg celastrol group). (C) The body weight and (D) the percent decrease (%) in body weight of the Lep−/− rats during the treatment period (n=5 for the vehicle group; n=6 for the 1,000 µg/kg celastrol group). The error bars represent the standard error of means. The P values were determined by one-way analysis of variance (ANOVA) or Student's t-test. aP<0.05.
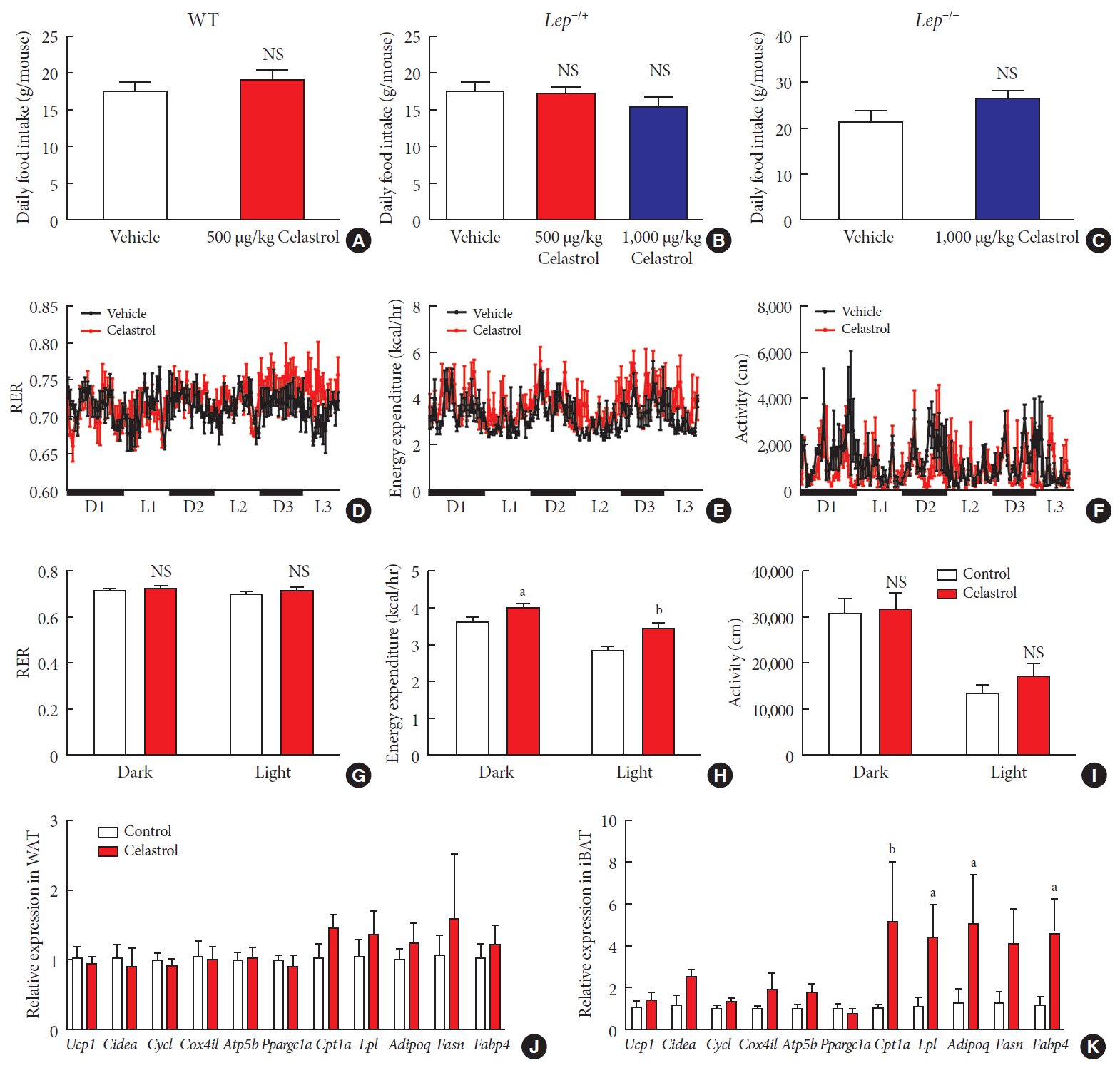
Fig. 5 Celastrol regulates energy homeostasis by enhancing energy expenditure (EE). (A–C) The average daily food intake of the (A) wild-type (WT), (B) Lep−/+, and (C) Lep−/− diet-induced obese (DIO) rats during the 3 weeks of treatment. (D-K) The WT DIO rats were placed in TSE Labmaster Caging System metabolic cages and administered vehicle or 500 µg/kg celastrol for 3 days after 1 week of celastrol acclimation (n=3 for each group). The dynamic or cycle-summarized (D, G) respiratory exchange ratios (RERs), (E, H) EE, and (F, I) the level of physical activity of each group of rats. (J, K) The mRNA expression of genes that participate in adipogenesis and fatty acid metabolism pathways in (J) white adipose tissue (WAT) and (K) brown adipose tissue (BAT). The error bars represent the standard error of means. (A-C and G-K) The P values were determined by Student's t-test. NS, not significant. aP<0.05, bP<0.01.
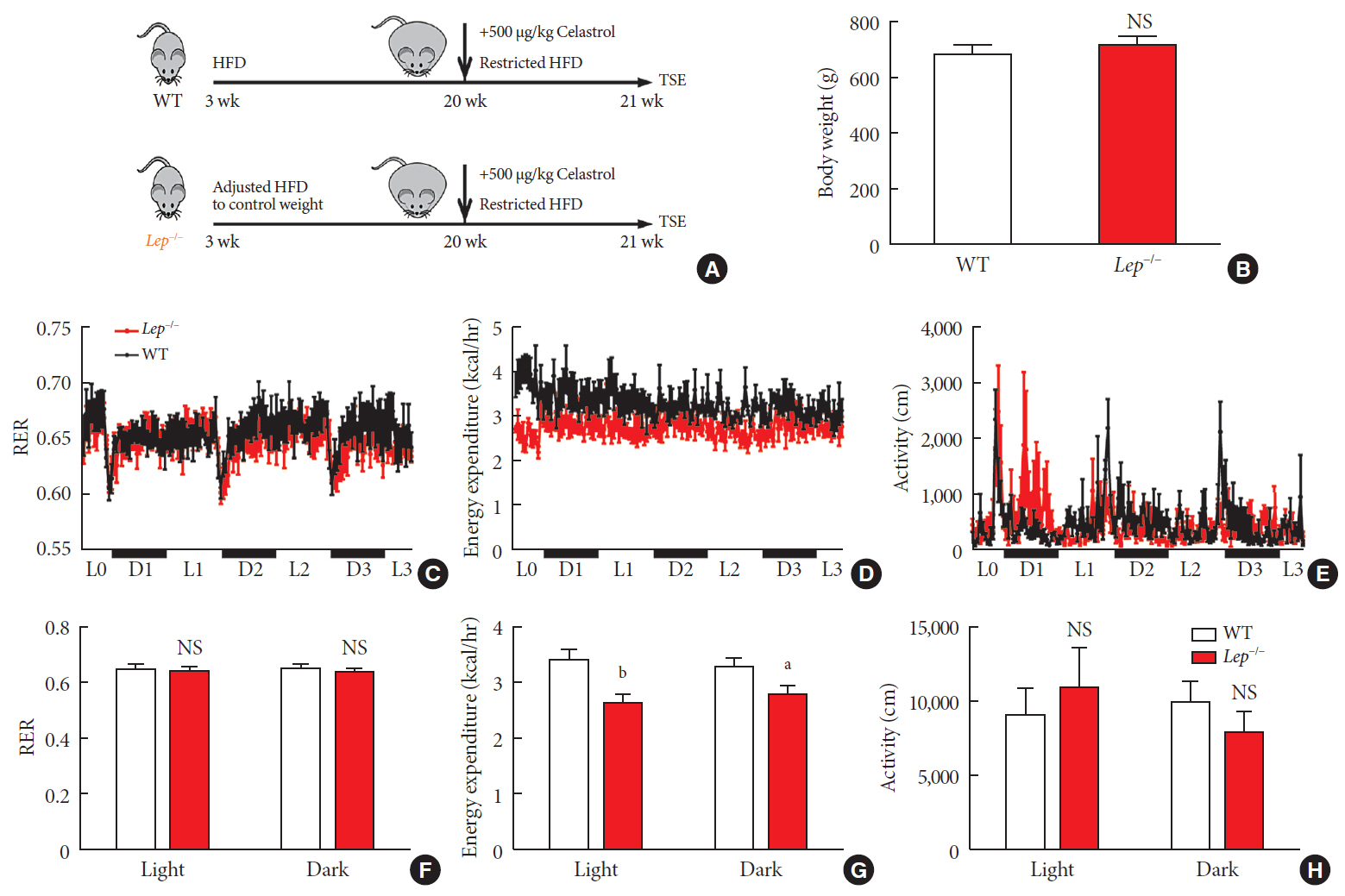
Fig. 6 Celastrol enhances energy expenditure (EE) via the leptin signaling pathway. To prevent the effects of different body weights on EE, wild-type (WT) and Lep−/− rats were fed a strictly controlled high-fat diet (HFD) to maintain equal body weights between the two groups. (A) A schematic diagram of diet control and (B) the body weight of the WT and Lep−/− rats before treatment with celastrol. (C–H) The WT and Lep−/− rats were placed in TSE Labmaster Caging System metabolic cages and administered vehicle or 500 µg/kg celastrol for 3 days after 1 week of celastrol acclimation (n=5 for each group). (C, F) The dynamic or cycle-summarized respiratory exchange ratios (RERs), (D, G) EE, and (E, H) the level of physical activity of each group of rats. The error bars represent the standard error of means. (B and F–H) The P values were determined by Student's t-test. NS, not significant. aP<0.05, bP<0.0001.
Reference
-
1. Yoshimoto S, Loo TM, Atarashi K, Kanda H, Sato S, Oyadomari S, Iwakura Y, Oshima K, Morita H, Hattori M, Honda K, Ishikawa Y, Hara E, Ohtani N. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome. Nature. 2013; 499:97–101.
Article2. Tseng YH, Cypess AM, Kahn CR. Cellular bioenergetics as a target for obesity therapy. Nat Rev Drug Discov. 2010; 9:465–482.
Article3. Al-Najim W, Docherty NG, le Roux CW. food intake and eating behavior after bariatric surgery. Physiol Rev. 2018; 98:1113–1141.
Article4. Cone RD. Studies on the physiological functions of the melanocortin system. Endocr Rev. 2006; 27:736–749.
Article5. Sohn JW, Elmquist JK, Williams KW. Neuronal circuits that regulate feeding behavior and metabolism. Trends Neurosci. 2013; 36:504–512.
Article6. Gropp E, Shanabrough M, Borok E, Xu AW, Janoschek R, Buch T, Plum L, Balthasar N, Hampel B, Waisman A, Barsh GS, Horvath TL, Bruning JC. Agouti-related peptide-expressing neurons are mandatory for feeding. Nat Neurosci. 2005; 8:1289–1291.
Article7. Paz-Filho G, Mastronardi CA, Licinio J. Leptin treatment: facts and expectations. Metabolism. 2015; 64:146–156.
Article8. Saltiel AR. New therapeutic approaches for the treatment of obesity. Sci Transl Med. 2016; 8:323rv2.
Article9. Chang CJ, Lin CS, Lu CC, Martel J, Ko YF, Ojcius DM, Tseng SF, Wu TR, Chen YY, Young JD, Lai HC. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat Commun. 2015; 6:7489.
Article10. Wu TR, Lin CS, Chang CJ, Lin TL, Martel J, Ko YF, Ojcius DM, Lu CC, Young JD, Lai HC. Gut commensal Parabacteroides goldsteinii plays a predominant role in the anti-obesity effects of polysaccharides isolated from Hirsutella sinensis. Gut. 2019; 68:248–262.
Article11. Zhang Z, Zhang H, Li B, Meng X, Wang J, Zhang Y, Yao S, Ma Q, Jin L, Yang J, Wang W, Ning G. Berberine activates thermogenesis in white and brown adipose tissue. Nat Commun. 2014; 5:5493.
Article12. Kannaiyan R, Shanmugam MK, Sethi G. Molecular targets of celastrol derived from Thunder of God Vine: potential role in the treatment of inflammatory disorders and cancer. Cancer Lett. 2011; 303:9–20.
Article13. Guo L, Luo S, Du Z, Zhou M, Li P, Fu Y, Sun X, Huang Y, Zhang Z. Targeted delivery of celastrol to mesangial cells is effective against mesangioproliferative glomerulonephritis. Nat Commun. 2017; 8:878.
Article14. Liu J, Lee J, Salazar Hernandez MA, Mazitschek R, Ozcan U. Treatment of obesity with celastrol. Cell. 2015; 161:999–1011.
Article15. Greenhill C. Celastrol identified as a leptin sensitizer and potential novel treatment for obesity. Nat Rev Endocrinol. 2015; 11:444.
Article16. Ma X, Xu L, Alberobello AT, Gavrilova O, Bagattin A, Skarulis M, Liu J, Finkel T, Mueller E. Celastrol protects against obesity and metabolic dysfunction through activation of a HSF1-PGC1α transcriptional axis. Cell Metab. 2015; 22:695–708.
Article17. Ridaura VK, Faith JJ, Rey FE, Cheng J, Duncan AE, Kau AL, Griffin NW, Lombard V, Henrissat B, Bain JR, Muehlbauer MJ, Ilkayeva O, Semenkovich CF, Funai K, Hayashi DK, Lyle BJ, Martini MC, Ursell LK, Clemente JC, Van Treuren W, Walters WA, Knight R, Newgard CB, Heath AC, Gordon JI. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science. 2013; 341:1241214.
Article18. Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006; 444:1027–1031.
Article19. Zhao L, Zhang F, Ding X, Wu G, Lam YY, Wang X, Fu H, Xue X, Lu C, Ma J, Yu L, Xu C, Ren Z, Xu Y, Xu S, Shen H, Zhu X, Shi Y, Shen Q, Dong W, Liu R, Ling Y, Zeng Y, Wang X, Zhang Q, Wang J, Wang L, Wu Y, Zeng B, Wei H, Zhang M, Peng Y, Zhang C. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science. 2018; 359:1151–1156.
Article20. Xu S, Zhu X, Li H, Hu Y, Zhou J, He D, Feng Y, Lu L, Du G, Hu Y, Liu T, Wang Z, Ding G, Chen J, Gao S, Wu F, Xue Z, Li Y, Fan G. The 14th Ile residue is essential for leptin function in regulating energy homeostasis in rat. Sci Rep. 2016; 6:28508.
Article21. Suárez-Zamorano N, Fabbiano S, Chevalier C, Stojanovic O, Colin DJ, Stevanovic A, Veyrat-Durebex C, Tarallo V, Rigo D, Germain S, Ilievska M, Montet X, Seimbille Y, Hapfelmeier S, Trajkovski M. Microbiota depletion promotes browning of white adipose tissue and reduces obesity. Nat Med. 2015; 21:1497–1501.
Article22. Callahan BJ, Sankaran K, Fukuyama JA, McMurdie PJ, Holmes SP. Bioconductor Workflow for Microbiome Data Analysis: from raw reads to community analyses. Version 2. F1000Res. 2016; 5:1492.23. Love MI, Anders S, Kim V, Huber W. RNA-seq workflow: gene-level exploratory analysis and differential expression. F1000Res. 2015; 4:1070.
Article24. Basso N, Soricelli E, Castagneto-Gissey L, Casella G, Albanese D, Fava F, Donati C, Tuohy K, Angelini G, La Neve F, Severino A, Kamvissi-Lorenz V, Birkenfeld AL, Bornstein S, Manco M, Mingrone G. Insulin resistance, microbiota, and fat distribution changes by a new model of vertical sleeve gastrectomy in obese rats. Diabetes. 2016; 65:2990–3001.
Article25. Turnbaugh PJ, Hamady M, Yatsunenko T, Cantarel BL, Duncan A, Ley RE, Sogin ML, Jones WJ, Roe BA, Affourtit JP, Egholm M, Henrissat B, Heath AC, Knight R, Gordon JI. A core gut microbiome in obese and lean twins. Nature. 2009; 457:480–484.
Article26. Cani PD, Bibiloni R, Knauf C, Waget A, Neyrinck AM, Delzenne NM, Burcelin R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes. 2008; 57:1470–1481.
Article27. Grasset E, Puel A, Charpentier J, Collet X, Christensen JE, Terce F, Burcelin R. A specific gut microbiota dysbiosis of type 2 diabetic mice induces GLP-1 resistance through an enteric NO-dependent and gut-brain axis mechanism. Cell Metab. 2017; 25:1075–1090.
Article28. Considine RV, Sinha MK, Heiman ML, Kriauciunas A, Stephens TW, Nyce MR, Ohannesian JP, Marco CC, McKee LJ, Bauer TL, Caro JF. Serum immunoreactive-leptin concentrations in normal-weight and obese humans. N Engl J Med. 1996; 334:292–295.
Article29. Ozcan U, Yilmaz E, Ozcan L, Furuhashi M, Vaillancourt E, Smith RO, Gorgun CZ, Hotamisligil GS. Chemical chaperones reduce ER stress and restore glucose homeostasis in a mouse model of type 2 diabetes. Science. 2006; 313:1137–1140.
Article30. Angulo MA, Butler MG, Cataletto ME. Prader-Willi syndrome: a review of clinical, genetic, and endocrine findings. J Endocrinol Invest. 2015; 38:1249–1263.
Article31. Bray GA, Fruhbeck G, Ryan DH, Wilding JP. Management of obesity. Lancet. 2016; 387:1947–1956.
Article32. Tremaroli V, Backhed F. Functional interactions between the gut microbiota and host metabolism. Nature. 2012; 489:242–249.
Article33. Zhao L. The gut microbiota and obesity: from correlation to causality. Nat Rev Microbiol. 2013; 11:639–647.
Article34. Martinez I, Stegen JC, Maldonado-Gomez MX, Eren AM, Siba PM, Greenhill AR, Walter J. The gut microbiota of rural papua new guineans: composition, diversity patterns, and ecological processes. Cell Rep. 2015; 11:527–538.35. Murphy EF, Cotter PD, Healy S, Marques TM, O'Sullivan O, Fouhy F, Clarke SF, O'Toole PW, Quigley EM, Stanton C, Ross PR, O'Doherty RM, Shanahan F. Composition and energy harvesting capacity of the gut microbiota: relationship to diet, obesity and time in mouse models. Gut. 2010; 59:1635–1642.
Article36. Ley RE, Turnbaugh PJ, Klein S, Gordon JI. Microbial ecology: human gut microbes associated with obesity. Nature. 2006; 444:1022–1023.37. Semova I, Carten JD, Stombaugh J, Mackey LC, Knight R, Farber SA, Rawls JF. Microbiota regulate intestinal absorption and metabolism of fatty acids in the zebrafish. Cell Host Microbe. 2012; 12:277–288.
Article38. Murphy EF, Cotter PD, Hogan A, O'Sullivan O, Joyce A, Fouhy F, Clarke SF, Marques TM, O'Toole PW, Stanton C, Quigley EM, Daly C, Ross PR, O'Doherty RM, Shanahan F. Divergent metabolic outcomes arising from targeted manipulation of the gut microbiota in diet-induced obesity. Gut. 2013; 62:220–226.
Article39. Hwang I, Park YJ, Kim YR, Kim YN, Ka S, Lee HY, Seong JK, Seok YJ, Kim JB. Alteration of gut microbiota by vancomycin and bacitracin improves insulin resistance via glucagon-like peptide 1 in diet-induced obesity. FASEB J. 2015; 29:2397–2411.40. Reijnders D, Goossens GH, Hermes GD, Neis EP, van der Beek CM, Most J, Holst JJ, Lenaerts K, Kootte RS, Nieuwdorp M, Groen AK, Olde Damink SW, Boekschoten MV, Smidt H, Zoetendal EG, Dejong CH, Blaak EE. Effects of gut microbiota manipulation by antibiotics on host metabolism in obese humans: a randomized double-blind placebo-controlled trial. Cell Metab. 2016; 24:63–74.
Article41. Hartstra AV, Bouter KE, Backhed F, Nieuwdorp M. Insights into the role of the microbiome in obesity and type 2 diabetes. Diabetes Care. 2015; 38:159–165.
Article42. Perry RJ, Peng L, Barry NA, Cline GW, Zhang D, Cardone RL, Petersen KF, Kibbey RG, Goodman AL, Shulman GI. Acetate mediates a microbiome-brain-β-cell axis to promote metabolic syndrome. Nature. 2016; 534:213–217.
Article43. Frederich RC, Hamann A, Anderson S, Lollmann B, Lowell BB, Flier JS. Leptin levels reflect body lipid content in mice: evidence for diet-induced resistance to leptin action. Nat Med. 1995; 1:1311–1314.
Article44. Kootte RS, Levin E, Salojarvi J, Smits LP, Hartstra AV, Udayappan SD, Hermes G, Bouter KE, Koopen AM, Holst JJ, Knop FK, Blaak EE, Zhao J, Smidt H, Harms AC, Hankemeijer T, Bergman JJGHM, Romijn HA, Schaap FG, Olde Damink SWM, Ackermans MT, Dallinga-Thie GM, Zoetendal E, de Vos WM, Serlie MJ, Stroes ESG, Groen AK, Nieuwdorp M. Improvement of insulin sensitivity after lean donor feces in metabolic syndrome is driven by baseline intestinal microbiota composition. Cell Metab. 2017; 26:611–619.
Article45. Everard A, Lazarevic V, Derrien M, Girard M, Muccioli GG, Neyrinck AM, Possemiers S, Van Holle A, Francois P, de Vos WM, Delzenne NM, Schrenzel J, Cani PD. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes. 2011; 60:2775–2786.
Article46. Leshan RL, Greenwald-Yarnell M, Patterson CM, Gonzalez IE, Myers MG Jr. Leptin action through hypothalamic nitric oxide synthase-1-expressing neurons controls energy balance. Nat Med. 2012; 18:820–823.
Article47. Pfuhlmann K, Schriever SC, Baumann P, Kabra DG, Harrison L, Mazibuko-Mbeje SE, Contreras RE, Kyriakou E, Simonds SE, Tiganis T, Cowley MA, Woods SC, Jastroch M, Clemmensen C, De Angelis M, Schramm KW, Sattler M, Messias AC, Tschop MH, Pfluger PT. Celastrol-induced weight loss is driven by hypophagia and independent from UCP1. Diabetes. 2018; 67:2456–2465.
Article48. Finan B, Yang B, Ottaway N, Smiley DL, Ma T, Clemmensen C, Chabenne J, Zhang L, Habegger KM, Fischer K, Campbell JE, Sandoval D, Seeley RJ, Bleicher K, Uhles S, Riboulet W, Funk J, Hertel C, Belli S, Sebokova E, Conde-Knape K, Konkar A, Drucker DJ, Gelfanov V, Pfluger PT, Muller TD, Perez-Tilve D, DiMarchi RD, Tschop MH. A rationally designed monomeric peptide triagonist corrects obesity and diabetes in rodents. Nat Med. 2015; 21:27–36.
Article
