Peanut sprout ethanol extract inhibits the adipocyte proliferation, differentiation, and matrix metalloproteinases activities in mouse fibroblast 3T3-L1 preadipocytes
- Affiliations
-
- 1Department of Food Science and Nutrition, Dankook University, 126 Jukjeon-dong, Suji-gu, Yongin-si, Gyeonggi 448-701, Korea. aewhaha@dankook.ac.kr
- 2Department of Food and Nutrition, Eulji University, Seongnam, Gyeonggi 461-723, Korea.
- 3Department of Food Engineering, Dankook University, Cheonan, Chungnam 330-714, Korea.
Abstract
- 3T3-L1 preadipocyte were differentiated to adipocytes, and then treated with 0, 10, 20, and 40 microg/mL of peanut sprout ethanol extract (PSEE). The main component of PSEE is resveratrol which contained 5.55 mg/mL of resveratrol. The MTT assay, Oil-Red O staining, glycerol-3-phosphate dehydrogenase (GPDH) activity, and the triglyceride concentration were determined in 3T3-L1 cells. MMP-2 and MMP-9 activities as well as mRNA expressions of C/EBP beta and C/EBP alpha were also investigated. As the concentration of PSEE in adipocytes increased, the cell proliferation was decreased in a dose-dependent manner from 4 days of incubation (P < 0.05). The GDPH activity (P < 0.05) and the triglyceride concentration (P < 0.05) were decreased as the PSEE treatment concentration increased. The mRNA expression of C/EBPbeta in 3T3-L1 cells was significantly low in groups of PSEE-treated, compared with control group (P < 0.05). The MMP-9 (P < 0.05) and MMP-2 (P < 0.05) activities were decreased in a dose-dependent manner as the PSEE concentration increased from 20 microg/mL. In conclusion, it was found that PSEE has an effect on restricting proliferation and differentiation of adipocytes.
Keyword
MeSH Terms
Figure
-
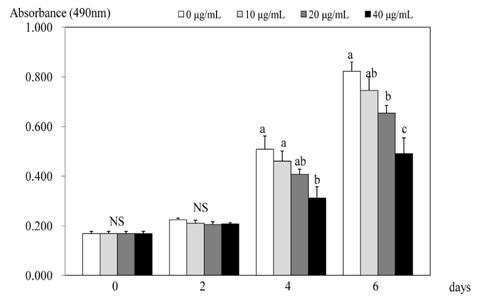
Fig. 1 Effect of PSEE on cell growth in 3T3-L1 cells. 3T3-L1 cells were plated at a density of 1.5 × 104 cells/mL in 24 well plate with DMEM, supplemented with 10% FBS, 10 µg/mL insulin, 1 µmol/L Dex, and 0.5 mmol/L IBMX for 2 days. After differentiation induction, the monolayer was incubated in post-differentiation medium with 0, 10, 20, or 40 µg/mL PSEE. Viable cell numbers were estimated by the MTT assay. Each bar represents the mean ± SE. Comparison among different concentrations of PSEE that yielded significant differences (P < 0.05) are indicated by different letters above each bar.
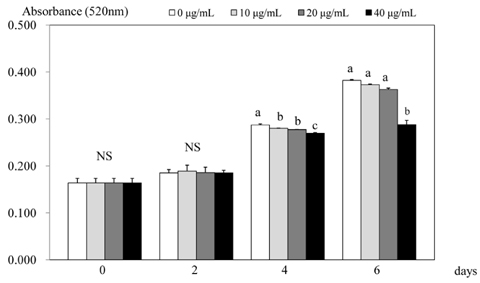
Fig. 2 Effect of PSEE on quantification of lipid content in 3T3-L1 cells. 3T3-L1 cells were plated at a density of 1.5 × 104 cells/mL in 24 well plate with DMEM supplemented with 10% FBS, 10 µg/mL insulin, 1 µmol/L Dex, and 0.5 mmol/L IBMX for 2 days. After differentiation induction, the monolayer was incubated in post-differentiation medium with 0, 10, 20, or 40 µg/mL PSEE. Lipid accumulation was estimated by the Oil-Red O staining. Each bar represents the mean ± SE. Comparison among different concentrations of PSEE that yielded significant differences (P < 0.05) are indicated by different letters above each bar.

Fig. 3 Effect of PSEE on triglyceride in 3T3-L1 cells. 3T3-L1 cells were plated at a density of 1.5 × 104 cells/mL in 24 well plate with DMEM supplemented with 10% FBS, 10 µg/mL insulin, 1 µmol/L Dex, and 0.5 mmol/L IBMX for 2 days. After differentiation induction, the monolayer was incubated in post-differentiation medium with 0, 10, 20, or 40 µg/mL PSEE. Triglyceride accumulation was estimated by the commercial triglyceride assay kit. Each bar represents the mean ± SE. Comparison among different concentrations of PSEE that yielded significant differences (P < 0.05) are indicated by different letters above each bar.
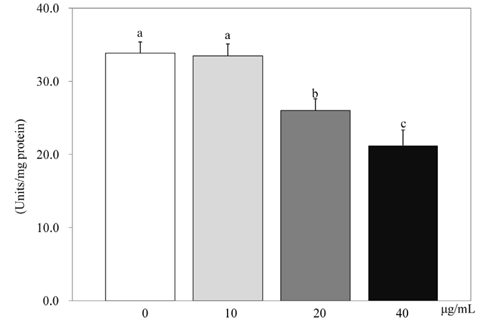
Fig. 4 Effect of PSEE on GPDH activity in 3T3-L1 cells. 3T3-L1 cells were plated at a density of 1.5 × 104 cells/mL in 24 well plate with DMEM supplemented with 10% FBS, 10 µg/mL insulin, 1 µmol/L Dex, and 0.5 mmol/L IBMX for 2 days. After differentiation induction, the monolayer was incubated in post-differentiation medium with 0, 10, 20, or 40 µg/mL PSEE. GPDH activity were estimated by commercial GPDH activity assay kit. Each bar represents the mean ± SE. Comparison among different concentrations of PSEE that yielded significant differences (P < 0.05) are indicated by different letters above each bar.
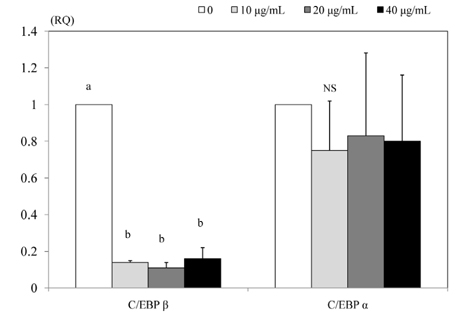
Fig. 5 Effect of PSEE on mRNA expression of transcription factors in 3T3-L1 cells. 3T3-L1 cells were plated at a density of 1.5 × 104 cells/mL in 24 well plate with DMEM supplemented with 10% FBS, 10 µg/mL insulin, 1 µmol/L Dex, and 0.5 mmol/L IBMX for 2 days. After differentiation induction, the monolayer was incubated in post-differentiation medium with 0, 10, 20, or 40 µg/mL PSEE. Chemiluminescent detection and quantitative analysis of western blots were performed for three independent experiments. The protein expression of C/EBP β (left) and C/EBP α (right) are shown above. Each bar represents the mean ± SE. Comparison among different concentrations of PSEE that yielded significant differences (P < 0.05) are indicated by different letters above each bar.

Fig. 6 Effect of PSEE on MMPs activity in 3T3-L1 cells. 3T3-L1 cells were plated at a density of 1.5 × 104 cells/mL in 24 well plate with DMEM supplemented with 10% FBS, 10 µg/mL insulin, 1 µmol/L Dex, and 0.5 mmol/L IBMX for 2 days. After differentiation induction, the monolayer was incubated in post-differentiation medium with 0, 10, 20, or 40 µg/mL PSEE. Medium were collected and concentrated for zymography. Above photograph of MMPs bands, which were representative of three independent experiments, are shown. Under bars were quantitative analyses of zymography. Each bar represents the mean ± SE. Comparison among different concentrations of PSEE that yielded significant differences (P < 0.05) are indicated by different letters above each bar.
Reference
-
1. Cornelius P, MacDougald OA, Lane MD. Regulation of adipocyte development. Annu Rev Nutr. 1994; 14:99–129.
Article2. Camp HS, Ren D, Leff T. Adipogenesis and fat-cell function in obesity and diabetes. Trends Mol Med. 2002; 8:442–447.
Article3. Rosen ED, Walkey CJ, Puigserver P, Spiegelman BM. Transcriptional regulation of adipogenesis. Genes Dev. 2000; 14:1293–1307.
Article4. Bouloumié A, Sengenès C, Portolan G, Galitzky J, Lafontan M. Adipocyte produces matrix metalloproteinases 2 and 9: involvement in adipose differentiation. Diabetes. 2001; 50:2080–2086.5. Bhat KP, Pezzuto JM. Cancer chemopreventive activity of resveratrol. Ann N Y Acad Sci. 2002; 957:210–229.
Article6. de la Lastra CA, Villegas I. Resveratrol as an antioxidant and pro-oxidant agent: mechanisms and clinical implications. Biochem Soc Trans. 2007; 35:1156–1160.
Article7. Das S, Alagappan VK, Bagchi D, Sharma HS, Maulik N, Das DK. Coordinated induction of iNOS-VEGF-KDR-eNOS after resveratrol consumption: a potential mechanism for resveratrol preconditioning of the heart. Vascul Pharmacol. 2005; 42:281–289.8. Kopp P. Resveratrol, a phytoestrogen found in red wine. A possible explanation for the conundrum of the 'French paradox'? Eur J Endocrinol. 1998; 138:619–620.
Article9. Hardie DG, Pan DA. Regulation of fatty acid synthesis and oxidation by the AMP-activated protein kinase. Biochem Soc Trans. 2002; 30:1064–1070.
Article10. Szkudelska K, Nogowski L, Szkudelski T. Resveratrol, a naturally occurring diphenolic compound, affects lipogenesis, lipolysis and the antilipolytic action of insulin in isolated rat adipocytes. J Steroid Biochem Mol Biol. 2009; 113:17–24.
Article11. Dal-Pan A, Blanc S, Aujard F. Resveratrol suppresses body mass gain in a seasonal non-human primate model of obesity. BMC Physiol. 2010; 10:11.
Article12. Gu X, Creasy L, Kester A, Zeece M. Capillary electrophoretic determination of resveratrol in wines. J Agric Food Chem. 1999; 47:3223–3227.
Article13. Wang KH, Lai YH, Chang JC, Ko TF, Shyu SL, Chiou RY. Germination of peanut kernels to enhance resveratrol biosynthesis and prepare sprouts as a functional vegetable. J Agric Food Chem. 2005; 53:242–246.
Article14. Kang HI, Kim JY, Park KW, Kang JS, Choi MR, Moon KD, Seo KI. Resveratrol content and nutritional components in peanut sprouts. Korean J Food Preserv. 2010; 17:384–390.15. Kang HI, Kim JY, Kwon SJ, Park KW, Kang JS, Seo KI. Antioxidative effects of peanut sprout extracts. J Korean Soc Food Sci Nutr. 2010; 39:941–946.
Article16. Lin BS, Lien TF, Chao MR, Lai TY, Chang JC, Chou SJ, Liao HF, Chiou RY. Toxicological and nutraceutical assessments of peanut sprouts as daily supplements to feed Sprague-Dawley rats for 18 weeks. J Sci Food Agric. 2008; 88:2201–2207.
Article17. Green H, Kehinde O. An established preadipose cell line and its differentiation in culture. II. Factors affecting the adipose conversion. Cell. 1975; 5:19–27.
Article18. Lee MS, Kim CT, Kim CJ, Cho YJ, Kim Y. Effects of Portulaca oleracea L. extract on lipolysis and hormone sensitive lipase (HSL) gene expression in 3T3-L1 adipocytes. Korean J Nutr. 2006; 39:742–747.19. Kim MJ, Kim Y, Chung JH, Kim JW, Kim HK. The effect of caffeine on 3T3-L1 adipocyte differentiation: a nutrigenomical approach. Korean J Nutr. 2005; 38:649–655.20. Chon JW, Sung JH, Hwang EJ, Park YK. Chlorella methanol extract reduces lipid accumulation in and increases the number of apoptotic 3T3-L1 cells. Ann N Y Acad Sci. 2009; 1171:183–189.
Article21. Tomiyama K, Nakata H, Sasa H, Arimura S, Nishio E, Watanabe Y. Wortmannin, a specific phosphatidylinositol 3-kinase inhibitor, inhibits adipocytic differentiation of 3T3-L1 cells. Biochem Biophys Res Commun. 1995; 212:263–269.
Article22. Ramírez-Zacarías JL, Castro-Muñozledo F, Kuri-Harcuch W. Quantitation of adipose conversion and triglycerides by staining intracytoplasmic lipids with Oil red O. Histochemistry. 1992; 97:493–497.
Article23. Na MH, Seo EY, Kim WK. Effects of α-lipoic acid on cell proliferation and apoptosis in MDA-MB-231 human breast cells. Nutr Res Pract. 2009; 3:265–271.
Article24. Kwon SY, Kang KJ. The effect of conjugated linoleic acid isomers on the cell proliferation, apoptosis and expressions of uncoupling protein (UCP) genes during differentiation of 3T3-L1 preadipocytes. Korean J Nutr. 2004; 37:533–539.25. Kyoya T, Ishida A, Nakashima K, Nakajima I, Toyoda A, Nakamura Y, Katsumata M. The effects of concentrations of lysine in media on differentiation of 3T3-L1 preadipocytes. Anim Sci J. 2011; 82:565–570.
Article26. Olofsson LE, Orho-Melander M, William-Olsson L, Sjöholm K, Sjöström L, Groop L, Carlsson B, Carlsson LM, Olsson B. CCAAT/enhancer binding protein alpha (C/EBPalpha) in adipose tissue regulates genes in lipid and glucose metabolism and a genetic variation in C/EBPalpha is associated with serum levels of triglycerides. J Clin Endocrinol Metab. 2008; 93:4880–4886.
Article27. Cho KJ, Moon HE, Moini H, Packer L, Yoon DY, Chung AS. Alpha-lipoic acid inhibits adipocyte differentiation by regulating pro-adipogenic transcription factors via mitogen-activated protein kinase pathways. J Biol Chem. 2003; 278:34823–34833.
Article28. Hong MK, Cho KY, Oh SJ, Kim KM, Yu SJ, Jung SS. Implications of the activation of matrix metalloproteinase-2 (MMP-2) on the metastasis in breast cancer. J Korean Surg Soc. 2002; 62:18–25.
Article29. Rural Development Administration, National Institute of Crop Science [Internet]. Suwon: Rural Development Administration;2010. cited 2010 October 14. Available from: http://rda.korea.kr/gonews.30. Shin HD. Anti-obesity effect of vegetable sprouts in 3T3-L1 adipocytes and rats [master's thesis]. Gwangju: Chosun University;2007.31. Wise LS, Green H. Participation of one isozyme of cytosolic glycerophosphate dehydrogenase in the adipose conversion of 3T3 cells. J Biol Chem. 1979; 254:273–275.
Article32. Rayalam S, Yang JY, Ambati S, Della-Fera MA, Baile CA. Resveratrol induces apoptosis and inhibits adipogenesis in 3T3-L1 adipocytes. Phytother Res. 2008; 22:1367–1371.
Article33. Zhang XH, Huang B, Choi SK, Seo JS. Anti-obesity effect of resveratrol-amplified grape skin extracts on 3T3-L1 adipocytes differentiation. Nutr Res Pract. 2012; 6:286–293.
Article34. Kang NE, Ha AW, Kim JY, Kim WK. Resveratrol inhibits the protein expression of transcription factors related adipocyte differentiation and the activity of matrix metalloproteinase in mouse fibroblast 3T3-L1 preadipocytes. Nutr Res Pract. 2012; 6:499–504.
Article35. Park HJ, Yang JY, Ambati S, Della-Fera MA, Hausman DB, Rayalam S, Baile CA. Combined effects of genistein, quercetin, and resveratrol in human and 3T3-L1 adipocytes. J Med Food. 2008; 11:773–783.
Article36. Folkman J, Klagsbrun M. Angiogenic factors. Science. 1987; 235:442–447.
Article37. Alexander CM, Selvarajan S, Mudgett J, Werb Z. Stromelysin-1 regulates adipogenesis during mammary gland involution. J Cell Biol. 2001; 152:693–703.
Article38. Gao D, Zhang X, Jiang X, Peng Y, Huang W, Cheng G, Song L. Resveratrol reduces the elevated level of MMP-9 induced by cerebral ischemia-reperfusion in mice. Life Sci. 2006; 78:2564–2570.
Article
- Full Text Links
-
- Actions
-
Cited
- CITED
-
- Close
- Share
-
- Similar articles
-
- Effects of quercetin on cell differentiation and adipogenesis in 3T3-L1 adipocytes
- Effects of (6)-gingerol, ginger component on adipocyte development and differentiation in 3T3-L1
- The Effects of Epigallocatechin on Adipogenesis of 3T3-L1 Preadipocytes
- Cryptotanshinone Inhibits Lipid Accumulation in Differentiating 3T3-L1 Preadipocytes by Down-regulating C/EBP-α, PPAR-γ, FAS, Perilipin A, and STAT-3
- Cryptotanshinone Inhibits Lipid Accumulation in Differentiating 3T3-L1 Preadipocytes by Down-regulating C/EBP-α, PPAR-γ, FAS, Perilipin A, and STAT-3
